 E-submission
E-submission
Articles
- Page Path
- HOME > J Pathol Transl Med > Volume 50(2); 2016 > Article
-
Original Article
Upregulated Neuro-oncological Ventral Antigen 1 (NOVA1) Expression Is Specific to Mature and Immature T- and NK-Cell Lymphomas - Eun Kyung Kim1, Sun Och Yoon1, Soo Hee Kim1,2, Woo Ick Yang1, Yoon Ah Cho1, Soo Jeong Kim3
-
Journal of Pathology and Translational Medicine 2016;50(2):104-112.
DOI: https://doi.org/10.4132/jptm.2016.02.08
Published online: February 29, 2016
1Department of Pathology, Yonsei University College of Medicine, Seoul, Korea
2Anatomic Pathology Reference Lab, Seegene Medical Foundation, Seoul, Korea
3Department of Internal Medicine, Division of Hematology, Yonsei University College of Medicine, Seoul, Korea
- Corresponding Author: Sun Och Yoon, MD, PhD Department of Pathology, Yonsei University College of Medicine, 50-1 Yonsei-ro, Seodaemun-gu, Seoul 03722, Korea Tel: +82-2-2228-1763 Fax: +82-2-362-0860 E-mail: soyoon@yuhs.ac
© 2016 The Korean Society of Pathologists/The Korean Society for Cytopathology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted noncommercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Background:
- Recent studies have revealed that the splicing factor neuro-oncological ventral antigen 1 (NOVA1) is enriched in fibroblasts and accumulated T cells of tertiary lymphoid structures. In the present study, we investigated NOVA1 expression in various subtypes of mature and immature T- and natural killer (NK)-cell lymphomas as well as in various B-cell lymphoma subtypes.
-
Methods:
- NOVA1 immunoexpression was evaluated in hyperplastic palatine tonsils (n = 20), T- and NK-cell lymphomas (n = 177), diffuse large B-cell lymphomas (n = 151), and other types of B cell lymphomas (n = 31). Nuclear staining intensity and percentage of positive tumor cells were graded. NOVA1 mRNA expression was analyzed in various lymphoma cell lines.
-
Results:
- Tumor cells of T- and NK-cell lymphomas showed higher expression levels of NOVA1 than did normal paracortical T cells, and 56.5% of T- and NK-cell lymphoma cases showed diffuse and strong expression. The NOVA1 expression level varied according to the subtype; it was higher in angioimmunoblastic T-cell lymphoma, anaplastic lymphoma kinase (ALK)-negative anaplastic large cell lymphoma (ALCL), and T lymphoblastic leukemia/lymphoma (T-LBL), but it was lower in ALK-positive ALCL. In almost all B-cell lymphomas, NOVA1 expression was very low or negative. NOVA1 mRNA was also expressed in Jurkat, a T-LBL cell line.
-
Conclusions:
- The present findings suggest that NOVA1 upregulation may be involved in certain subtypes of T- and NK-cell lymphomas, but not in B-cell lymphomas. Upregulated NOVA1 expression seems to be a specific biological feature of activated T cells such as T- and NK-cell lymphomas.
- Patients and clinical data
- Archival tissues of formalin-fixed, paraffin-embedded (FFPE) tumor blocks were retrieved from consecutively archived specimens. A set of 177 T/NK-cell lymphomas diagnosed in Severance Hospital from 1999 to 2013 were included. The subtypes of T/NK-cell lymphomas were as follows: extranodal NK/T-cell lymphoma, nasal type (NKTL; n=60), peripheral T-cell lymphoma, not otherwise specified (PTCL-NOS; n=44), angioimmunoblastic T-cell lymphoma (AITL; n=16), anaplastic large cell lymphoma (ALCL; anaplastic lymphoma kinase [ALK]-positive, n=16; ALK-negative, n=17), T lymphoblastic leukemia/lymphoma (T-LBL; n=20) and other types (n=4; hepatosplenic T-cell lymphoma, n=2; subcutaneous panniculitis-like T-cell lymphoma, n=1; cutaneous gamma-delta T-cell lymphoma, n=1). Clinical information was obtained from patient medical records and 139 had clinical data available. The clinicopathological characteristics are summarized in Appendix 1.
- A set of 151 diffuse large B cell lymphoma (DLBCL) and other types of 31 B-cell lymphomas, which were diagnosed in Severance Hospital from 2005 to 2011, were included. Other types of B-cell lymphomas were as follows: follicular lymphoma (n=14), mantle cell lymphoma (n=5), marginal zone B-cell lymphoma (n=7), and Burkitt lymphoma (n=5). Hyperplastic palatine tonsil tissues of cancer-free individuals (n=20) were randomly retrieved. All tumor samples were reviewed by hematopathologists based on the current World Health Organization (WHO) criteria [11]. The institutional review board approved this study.
- Tissue microarray preparation and immunohistochemistry
- Sections of FFPE tissues were prepared and stained with hematoxylin and eosin. Under the microscope, the representative area was confirmed and selected for a 3-mm-core sized tissue microarray (TMA). One or two different representative areas per case were selected and 29 cores were embedded in a TMA block.
- Immunohistochemistry of NOVA1 (1:500, Abcam, Cambridge, UK) was performed on 4-μm tissue sections using the Ventana BenchMark XT Autostainer (Ventana Medical Systems, Tucson, AZ, USA). Immunohistochemistry with other primary antibodies was performed for diagnostic purposes and the tested antibodies are listed in Appendix 2.
- Analysis of immunohistochemistry
- NOVA1 expression was semiquantitatively determined according to the H-score method with a total score range of 0–300 [12]. The dominant intensity score of nuclear staining (0, absent; 1, weak or barely detectable; 2, distinct brown; and 3, strong dark brown) was multiplied by the percentage of positive tumor cell nuclei. The NOVA1 protein expression score ranged from 0 to 300, as high (200<H-score≤300), intermediate (100<H-score ≤200), or low (0≤H-score≤100) expression.
- Cell lines and reagents
- Jurkat, a T-LBL cell line, was maintained in RPMI1640 medium (22400-089, Gibco, Life Technologies, Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; 6000-044, Gibco, Life Technologies). HH, a cell line derived from CD30-positive cutaneous T-cell lymphoma, and MAC1, a cell line derived from CD30-positive ALK-negative ALCL, were maintained in RPMI1640 medium supplemented with 20% heat-inactivated FBS. SNK6, derived from Epstein-Barr virus (EBV)-positive NKTL, was cultured in RPMI1640 medium supplemented with 10% heat-inactivated human plasma and 700 U/mL of recombinant interleukin 2. YT, an EBV-positive human NK-like leukemic cell line, was cultured in Iscove’s modified Dulbecco’s medium (12440-046, Gibco) supplemented with 20% heat-inactivated FBS. All the cell lines derived from T- or NK-cell lymphomas were kindly provided by Prof. Jeon YK (Seoul National University College of Medicine, Seoul, Korea). Toledo, a DLBCL cell line, was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) and maintained in RPMI1640 supplemented with 10% heat-inactivated FBS.
- RNA isolation and quantitative polymerase chain reaction
- RNA isolation was performed using an RNeasy plus mini kit (74134, Qiagen, Hilden, Germany), and cDNA was constructed using a cDNA synthesis kit (11754-050, Invitrogen, Carlsbad, CA, USA). The TaqMan probes used were as follows: GAPDH (Hs99999905-m1, AB, Life Technologies) and NOVA1 (Hs00359592-m1, AB, Life Technologies). Quantitative polymerase chain reaction was performed using ABI StepOnePlus under the following conditions: 50°C for 2 minutes, 95°C for 5 minutes, 40 cycles of 95°C for 15 seconds, and 60°C for minutes. Relative mRNA expression levels of NOVA1 were determined by the comparative method (2-ddCt) against the reference GAPDH.
- Statistical analysis
- The Kruskal-Wallis (K-W), Mann-Whitney U (M-W), t and χ2 tests were used to analyze differences between the variables examined. Survival rates were analyzed with the Kaplan-Meier method, and differences were compared using the log-rank test. Two-sided p-values less than .05 were considered statistically significant. Statistical analyses were done using the IBM SPSS ver. 22 (IBM Corp., Armonk, NY, USA).
MATERIALS AND METHODS
- Expression of NOVA1 in the palatine tonsils
- The NOVA1 expression patterns in the cell nuclei were similar to those reported in a previous study [6]. Stromal spindle cells/fibroblasts showed strong and constant expression of NOVA1. Endothelial cells variably expressed NOVA1 with negative to moderate intensity. Almost all B cells of the germinal centers or outside the germinal centers were negative for NOVA1. As for T cells, occasional expression of NOVA1 (intensity grade 2–3 with 5% or less percentage of T cells) was noted in the paracortical area (Table 1).
- Expression of NOVA1 in T/NK-cell lymphomas
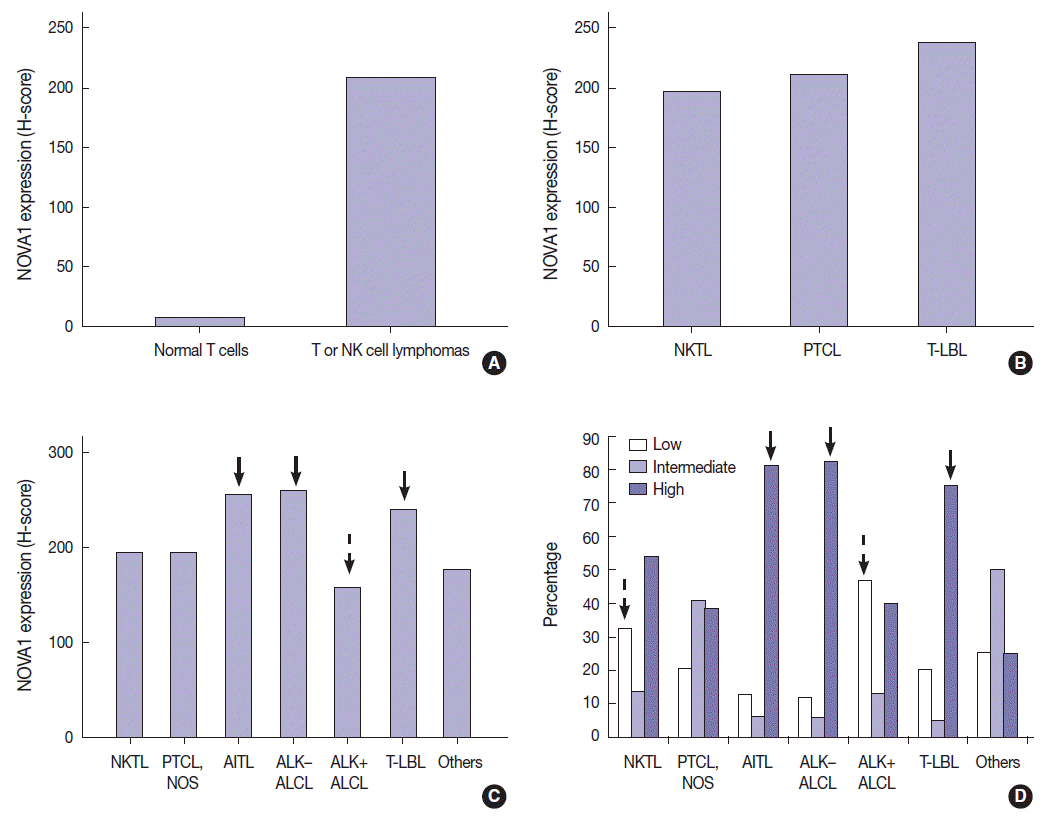
- T/NK-cell lymphomas showed low to high NOVA1 expression and 99 (56.5%) showed high NOVA1 expression (Table 1, Fig. 1). The expression level (score) of NOVA1 was higher in tumor cells of T/NK-cell lymphomas when compared to normal T cells of tonsil tissues (mean score, 209 vs. 7; p<.001; non-parametric M-W) (Fig. 2A). Within T/NK-cell lymphomas, precursor T-LBL showed a relatively higher level of NOVA1 expression than did NKTL or mature (peripheral) T-cell lymphomas, although a statistically significant difference was not noted (p=.226; K-W) (Fig. 2B). According to the specific subtypes of T/NK-cell lymphomas, the expression level of NOVA1 in tumor cells varied (p=.011; K-W). AITL, ALK-negative ALCL, and T-LBL showed relatively higher NOVA1 expression levels when compared to other subtypes (Table 1, Fig. 2C). Specifically, ALK-positive ALCL revealed a lower level of NOVA1 expression when compared to ALK-negative ALCL (mean score, 156 vs. 259; p=.027; M-W) and ALK-positive ALCL showed the lowest expression level among subtypes of T/NK-cell lymphomas (Table 1, Fig. 2C). The rate of high, intermediate, or low expression of NOVA1 was also different according to subtype (p <.001); high expression was relatively more frequent in AITL, ALK-negative ALCL, and T-LBL cells, while low expression was relatively more frequent in NKTL and ALK-positive ALCL (Table 1, Fig. 2D).
- Expression of NOVA1 in DLBCLs
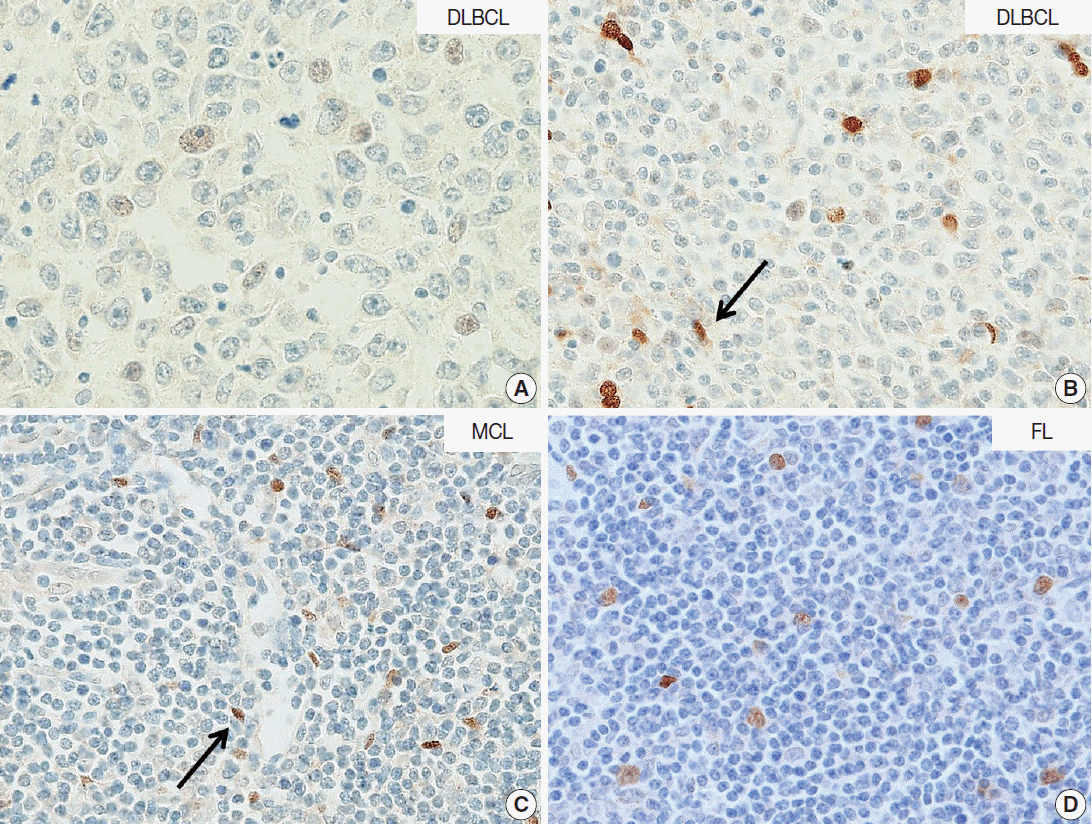
- In DLBCL, high NOVA1 expression was not noted in any cases. The tested 151 DLBCL cases revealed an expression pattern with a score less than 10. Furthermore, in the various types of other B-cell lymphomas, high NOVA1 expression was not noted (Table 1, Fig. 3).
- NOVA1 gene expression in human lymphoma cell lines
- NOVA1 mRNA was expressed only in the Jurkat cell line, which corresponds to T-LBL. NOVA1 mRNA was not detected in other tested cell lines of HH (correspondent to peripheral, mature T cell lymphoma), MAC1 (peripheral, mature T-cell lymphoma), SNK6 (NKTL), YT (NK cell leukemia), or Toledo (DLBCL) (Fig. 4).
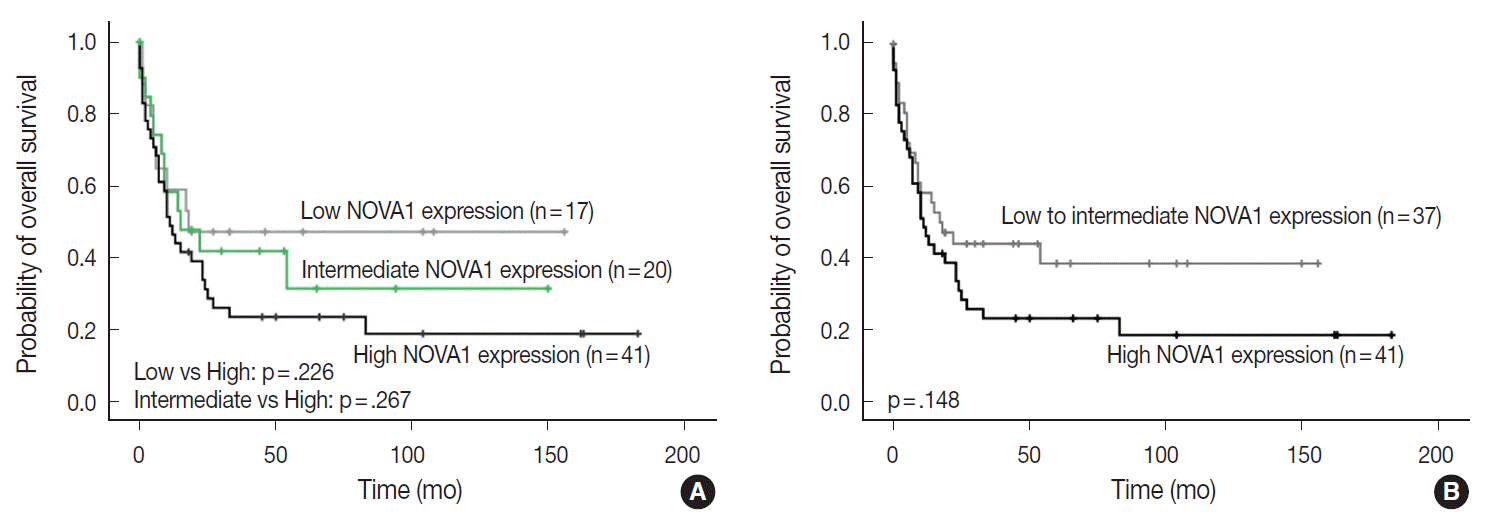
- NOVA1 expression and its clinicopathologic correlation in T/NK-cell lymphomas
- The NOVA1 expression level in tumor cells was not different according to the organ/sites where lymphomas were growing (Appendix 3). The known clinicopathological prognostic factors such as age, lactate dehydrogenase level, Ann-Arbor stage, or International Prognostic Index (IPI) risk scores showed no correlation with NOVA1 expression. In cases of mature (peripheral) T-cell lymphomas, high expression of NOVA1 tended to be related to shorter overall survival rate in comparison to low or intermediate NOVA1 expression, although statistical significance was not observed (Fig. 5A, B). In other subtypes, such as NKTL and T-LBL, no correlation was noted between NOVA1 expression and overall survival rate.
RESULTS
- In this study, we found that NOVA1 was frequently and highly expressed in various subtypes of T/NK-cell lymphomas, with about 60% of T/NK-cell lymphoma cases showing high NOVA1 expression, whereas only a small number of T cells expressed NOVA1 in the hyperplastic palatine tonsils. As for B-cell lymphomas, NOVA1 was negative in various types of B-cell lymphomas other than DLBCL, or normal B cells. In DLBCL, NOVA1 was expressed in a few cases; however, high expression was not noted in any cases.
- Our previous study [6] revealed high expression of NOVA1 in activated T cells and the present study also confirmed that finding. Moreover, NOVA1 was upregulated in lymphomas derived from immature T cells such as precursor T-LBL and NKTL. However, in contrast to T/NK cells, NOVA1 might not be involved in B cell lesions, either malignant lymphomas or non-neoplastic B cells, as shown in our studies.
- Among subtypes of T/NK-cell lymphomas, the expression level in tumor cells was different. Regarding the different biological characteristics and pathogenesis of each subtype of T/NK-cell lymphoma, NOVA1 expression may be affected by various intrinsic biological factors related to each subtype. The AITL, ALK-negative ALCL, and T-LBL subtypes, which showed distinctively high NOVA1 expression, are biologically characteristic entities [11]. Interestingly, the NOVA1 expression pattern was different even within ALCL specimens, according to ALK expression status. ALK-negative ALCL showed distinctively high NOVA1 expression while ALK-positive ALCL exhibited low NOVA1 expression. Both types of ALCL share common morphologic features of CD30 positivity, large anaplastic “hallmark” cells, frequent loss of pan-T-cell antigens, and expression of cytotoxic molecules, and are classified as the same disease. However, ALK-positive ALCL and ALK-negative ALCL are thought to be different entities; the clinical features, prognosis, and gene expression signatures have been noted to be different [11] Therefore, the differences in NOVA1 expression may be related to the intrinsic biologic characteristics of these two types. There are no known studies that can explain the distinctively low NOVA1 expression in ALK-positive ALCL. We thought that nucleophosmin (NPM), the fusion partner of ALK-NPM translocation/t(2;5) in ALK-positive ALCLs, may be associated. NPM is a putative ribosome assembly factor involved in processing preRNA molecules [13]. Mutated NPM molecules are abnormally localized in the cytoplasm because of impaired nuclear localization, and this abnormal localization of NPM is thought to be associated with abnormal ribosome biogenesis [13]. The aberrant function of mutated NPM may be related to inhibiting NOVA1 function in the splicing process of pre-RNA molecules via abnormal ribosome biogenesis. PTCL-NOS showed a relatively diverse pattern of NOVA1 expression. The proportion of cases showing an intermediate level of NOVA1 expression was relatively higher than that in other subtypes. This difference may be because PTCL-NOS is a disease group of heterogeneous entities of mature (peripheral) T-cell lymphomas.
- NOVA1 expression did not vary according to the organ site where the lymphoma developed. These findings indicate that NOVA1 expression may be primarily dependent on the intrinsic characteristics of the tumor cell itself rather than extrinsic factors such as the organ-specific microenvironment, which is pre-formed before tumor growth. In association with patient prognosis, NOVA1 expression showed no statistical significance. However, in mature (peripheral) T-cell lymphomas, high NOVA1 expression tended to be related to an inferior overall survival rate. In our previous study, high NOVA1 expression levels in reactive, non-neoplastic (mature) T cells, which accumulate within the tertiary lymphoid structures of gastric carcinoma microenvironment, were associated with favorable patient prognosis. Suppression of NOVA1 in such reactive T cells was related with poor patient prognosis, and this effect is thought to be related to the suppression of regulatory T cell functions [6]. From these findings, the role of NOVA1 in tumor T cells of malignant lymphomas and non-tumor T cells of tissues seems to be different. In normal physiologic conditions, NOVA1 upregulation seems to be involved in normal immune regulation. In neoplastic T cells, however, NOVA1 upregulation seems to be involved in tumor progression.
- Alternative splicing of pre-mRNA is the main mechanism of transcriptome and proteome diversity [4,14], and the splicing program relies on the action of splicing factors [1] like NOVA1. The action of splicing factors is cell type-specific and independent of organ origin [2,3,5]. Previous studies have suggested or shown that normal and tumorous hematopoietic cells are regulated by specific alternative splicing programs and alternative transcript variants such as c-myb, a transcription factor of hematopoietic cells; CD45, a transmembrane protein tyrosine phosphatase of T cells, B cells, and other hematopoietic cells; and CD20, a surface marker of mature B cells [15-17]. Likewise, NOVA1 may work as an alternative splicing factor in the lymphoreticular system and in lymphomagenesis of T/NK-cell lineage.
- Among the tested cell lines, only Jurkat cells, which corresponded to immature T cells, expressed the NOVA1 gene. These findings could partially support the findings observed in clinical patient samples. The functional role of NOVA1 in lymphoid cells and target genes regulated by NOVA1 in lymphomagenesis should be identified through further studies.
- In summary, NOVA1 was frequently upregulated in various subtypes of T/NK-cell lymphomas. NOVA1 expression may be a new biomarker for T/NK-cell lymphomas and should be studied as a possible therapeutic target for this type of lymphoma.
DISCUSSION
Acknowledgments
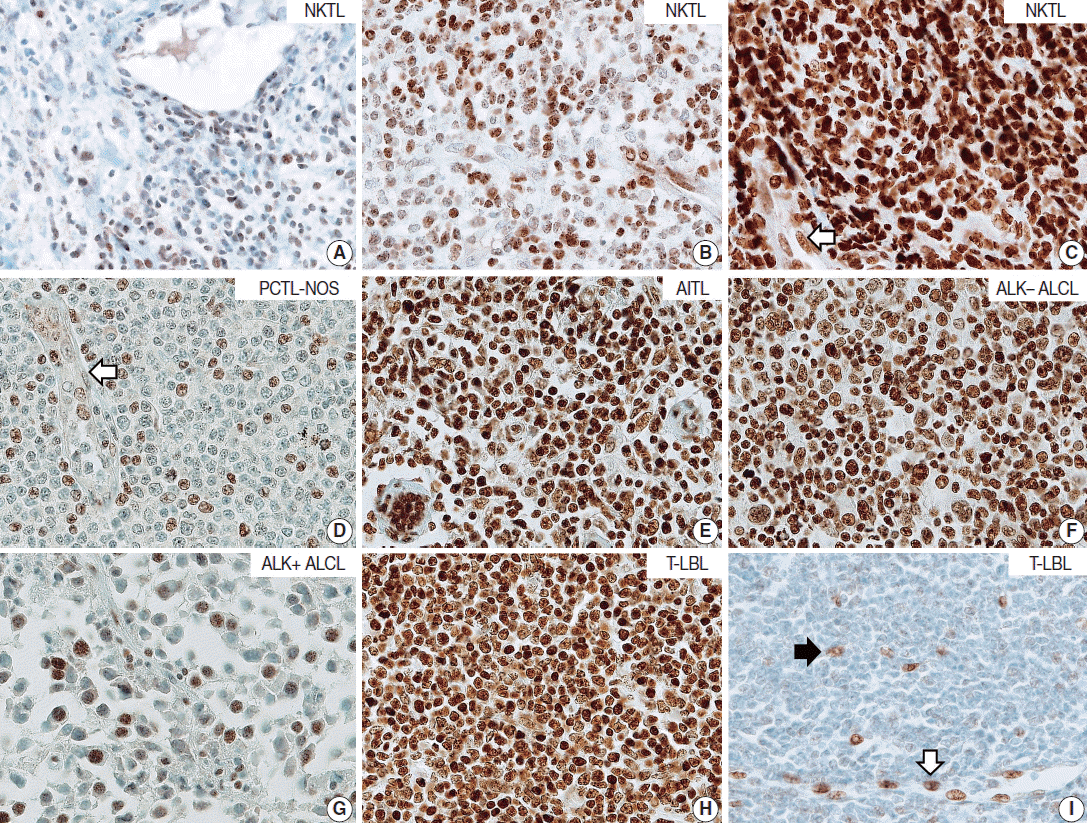
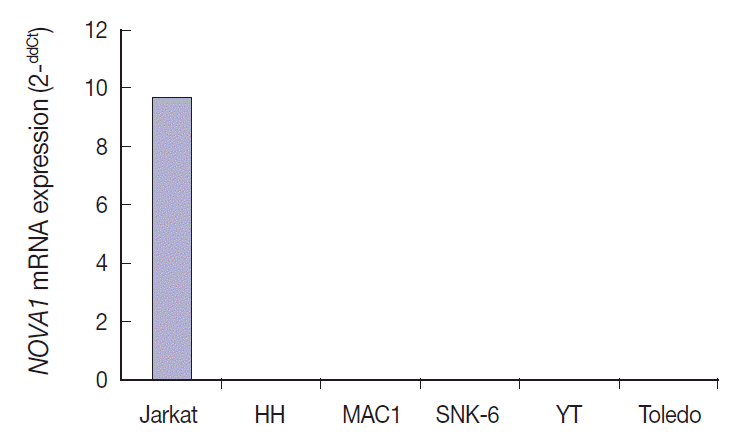
| Tissue category | Case No. |
NOVA1 expression |
||||
|---|---|---|---|---|---|---|
| General pattern |
Frequency of expression category (%) |
H-score (mean) | ||||
| High | Intermediate | Low | ||||
| Palatine tonsils | 20 | Small numbers of T cells | 0 | 0 | 100 | 7 |
| T- and NK-cell lymphomas | 177 | Strong and diffuse | 56.5 | 18.6 | 24.9 | 209 |
| NKTL | 60 | Lower | 54.2 | 13.6 | 32.2 | 195 |
| PTCL-NOS | 44 | Diverse | 38.6 | 40.9 | 20.5 | 194 |
| AITL | 16 | Higher | 81.2 | 6.2 | 12.5 | 255 |
| ALK-negative ALCL | 17 | Higher | 82.4 | 5.9 | 11.8 | 259 |
| ALK-positive ALCL | 16 | Lower | 40 | 13.3 | 46.7 | 156 |
| T-LBL | 20 | Higher | 75 | 5 | 20 | 237 |
| Other typesa | 4 | NA | 25 | 50 | 25 | 175 |
| B-Cell lymphomas | 182 | Almost negative | 0 | 0 | 100 | 7 |
| DLBCL | 151 | Almost negative | 0 | 0 | 100 | 8 |
| Other typesb | 31 | Almost negative | 0 | 0 | 100 | 5 |
NOVA1, neuro-oncological ventral antigen 1; NK, natural killer; NKTL, extranodal natural killer/T-cell lymphoma, nasal type; PTCL-NOS, peripheral T-cell lymphoma, not otherwise specified; AITL, angioimmunoblastic T-cell lymphoma; ALK, anaplastic lymphoma kinase; ALCL, anaplastic large cell lymphoma; T-LBL, T lymphoblastic leukemia/lymphoma; DLBCL, diffuse large B-cell lymphoma.
aThe T- or NK-cell lymphomas of the “other type” category included hepatosplenic T-cell lymphoma (n = 2), subcutaneous panniculitis-like T-cell lymphoma (n = 1), and cutaneous gamma-delta T-cell lymphoma (n = 1). The case number for this category was too small (only 4 cases), and therefore NOVA1 expression may not be representative for this category;
bThe set of “other type” B-cell lymphomas was composed of follicular lymphoma (n = 14), mantle cell lymphoma (n = 5), marginal zone B-cell lymphoma (n = 7), and Burkitt lymphoma (n = 5).
- 1. Barash Y, Calarco JA, Gao W, et al. Deciphering the splicing code. Nature 2010; 465: 53-9. ArticlePubMedPDF
- 2. de la Grange P, Gratadou L, Delord M, Dutertre M, Auboeuf D. Splicing factor and exon profiling across human tissues. Nucleic Acids Res 2010; 38: 2825-38. ArticlePubMedPMC
- 3. Merkin J, Russell C, Chen P, Burge CB. Evolutionary dynamics of gene and isoform regulation in mammalian tissues. Science 2012; 338: 1593-9. ArticlePubMedPMC
- 4. Kalsotra A, Cooper TA. Functional consequences of developmentally regulated alternative splicing. Nat Rev Genet 2011; 12: 715-29. ArticlePubMedPMCPDF
- 5. Mallinjoud P, Villemin JP, Mortada H, et al. Endothelial, epithelial, and fibroblast cells exhibit specific splicing programs independently of their tissue of origin. Genome Res 2014; 24: 511-21. ArticlePubMedPMC
- 6. Yoon SO, Kim EK, Lee M, et al. NOVA1 inhibition by miR-146b-5p in the remnant tissue microenvironment defines occult residual disease after gastric cancer removal. Oncotarget 2016; 7: 2475-95. ArticlePubMedPMC
- 7. Drayton DL, Liao S, Mounzer RH, Ruddle NH. Lymphoid organ development: from ontogeny to neogenesis. Nat Immunol 2006; 7: 344-53. ArticlePubMedPDF
- 8. Ruddle NH, Akirav EM. Secondary lymphoid organs: responding to genetic and environmental cues in ontogeny and the immune response. J Immunol 2009; 183: 2205-12. ArticlePubMedPMCPDF
- 9. Buckley CD, Barone F, Nayar S, Bénézech C, Caamaño J. Stromal cells in chronic inflammation and tertiary lymphoid organ formation. Annu Rev Immunol 2015; 33: 715-45. ArticlePubMed
- 10. Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer 2006; 6: 392-401. ArticlePubMedPDF
- 11. Swerdlow SH, Campo E, Harris NL, et al. WHO classification of tumours of haematopoietic and lymphoid tissues. 4th ed. Lyon: IARC Press, 2008.
- 12. Park E, Park SY, Kim H, et al. Membranous insulin-like growth factor-1 receptor (IGF1R) expression is predictive of poor prognosis in patients with epidermal growth factor receptor (EGFR)-mutant lung adenocarcinoma. J Pathol Transl Med 2015; 49: 382-8. ArticlePubMedPMCPDF
- 13. Falini B, Nicoletti I, Bolli N, et al. Translocations and mutations involving the nucleophosmin (NPM1) gene in lymphomas and leukemias. Haematologica 2007; 92: 519-32. ArticlePubMed
- 14. Kelemen O, Convertini P, Zhang Z, et al. Function of alternative splicing. Gene 2013; 514: 1-30. ArticlePubMed
- 15. Oberdoerffer S, Moita LF, Neems D, Freitas RP, Hacohen N, Rao A. Regulation of CD45 alternative splicing by heterogeneous ribonucleoprotein, hnRNPLL. Science 2008; 321: 686-91. ArticlePubMedPMC
- 16. O’Rourke JP, Ness SA. Alternative RNA splicing produces multiple forms of c-Myb with unique transcriptional activities. Mol Cell Biol 2008; 28: 2091-101. ArticlePubMedPMCPDF
- 17. Henry C, Deschamps M, Rohrlich PS, et al. Identification of an alternative CD20 transcript variant in B-cell malignancies coding for a novel protein associated to rituximab resistance. Blood 2010; 115: 2420-9. ArticlePubMedPDF
REFERENCES
Appendix
Appendix 1.
NK, natural killer; NKTL, extranodal natural killer/T-cell lymphoma, nasal type; PTCL, peripheral T-cell lymphoma; AITL, angioimmunoblastic T-cell lymphoma; ALCL, anaplastic large cell lymphoma; ALK, anaplastic lymphoma kinase; T-LBL, T lymphoblastic leukemia/lymphoma; H&N, head and neck; LN, lymph node; GI, gastrointestinal tract; LDH, lactate dehydrogenase; BM, bone marrow; IPI, International Prognostic Index; CTx, chemotherapy; RTx, radiotherapy; Tx, treatment.
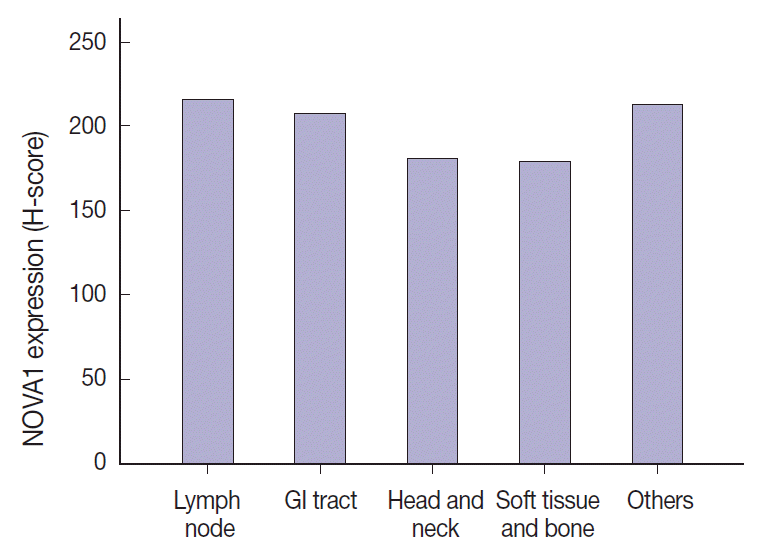
Figure & Data
References
Citations
- Intraoperative pathologic diagnosis of central nervous system lymphomas: A comparison of frozen and permanent section diagnoses, and the significance of preoperative imaging
Aslı Kahraman, Fikret Dirilenoğlu, İsmail Güzeliş, Kenan Çetinoğlu
Annals of Diagnostic Pathology.2024; 69: 152246. CrossRef - NOVA1 acts on Impact to regulate hypothalamic function and translation in inhibitory neurons
Yoko Tajima, Keiichi Ito, Yuan Yuan, Mayu O. Frank, Yuhki Saito, Robert B. Darnell
Cell Reports.2023; 42(2): 112050. CrossRef - MicroRNA miR-27a-3p accelerates cardiac hypertrophy by targeting neuro-oncological ventral antigen 1
Dongyun Li, Mingzhi Shen, Xinxin Deng, Yongyi Bai
Bioengineered.2022; 13(4): 8982. CrossRef - NOVA1 promotes NSCLC proliferation and invasion by activating Wnt/β-catenin signaling
Lianyue Qu, Yulong Tian, Fan Wang, Zixuan Li
BMC Cancer.2022;[Epub] CrossRef - Identification of the Functions and Prognostic Values of RNA Binding Proteins in Bladder Cancer
Yue Wu, Zheng Liu, Xian Wei, Huan Feng, Bintao Hu, Bo Liu, Yang Luan, Yajun Ruan, Xiaming Liu, Zhuo Liu, Shaogang Wang, Jihong Liu, Tao Wang
Frontiers in Genetics.2021;[Epub] CrossRef - Nervous system and gastric cancer
Ke Wang, Xin-hui Zhao, Jun Liu, Rui Zhang, Ji-peng Li
Biochimica et Biophysica Acta (BBA) - Reviews on Cancer.2020; 1873(1): 188313. CrossRef - The RNA-binding protein RBM47 inhibits non-small cell lung carcinoma metastasis through modulation of AXIN1 mRNA stability and Wnt/β-catentin signaling
Di-jian Shen, You-hua Jiang, Jian-qiang Li, Li-wei Xu, Kai-yi Tao
Surgical Oncology.2020; 34: 31. CrossRef - Genome-Wide Profiling of Cervical RNA-Binding Proteins Identifies Human Papillomavirus Regulation of RNASEH2A Expression by Viral E7 and E2F1
Junfen Xu, Habin Liu, Yanqin Yang, Xiaohong Wang, Poching Liu, Yang Li, Craig Meyers, Nilam Sanjib Banerjee, Hsu-Kun Wang, Maggie Cam, Weiguo Lu, Louise T. Chow, Xing Xie, Jun Zhu, Zhi-Ming Zheng, Xiang-Jin Meng, James Pipas, Xuefeng Liu, Lucia Pirisi-cre
mBio.2019;[Epub] CrossRef - NOVA1 induction by inflammation and NOVA1 suppression by epigenetic regulation in head and neck squamous cell carcinoma
Eun Kyung Kim, Yoon Ah Cho, Mi-kyoung Seo, Hyunmi Ryu, Byoung Chul Cho, Yoon Woo Koh, Sun Och Yoon
Scientific Reports.2019;[Epub] CrossRef - The RNA binding protein neuro‐oncological ventral antigen 1 (NOVA1) regulates IL-6 mRNA stability to enhance JAK2-STAT3 signaling in CRC
Yong-gang Hong, Guo-shu Xu, Guan-yu Yu, Ji-dian Zhou, Qi-zhi Liu, Jun-sheng Ni, Hong-li Yan, Wei Zhang, Li-qiang Hao
Surgical Oncology.2019; 31: 67. CrossRef - NOVA1 acts as an oncogene in melanoma via regulating FOXO3a expression
Xin Yu, Heyi Zheng, Matthew T.V. Chan, William K.K. Wu
Journal of Cellular and Molecular Medicine.2018; 22(5): 2622. CrossRef - Neuro‐oncological ventral antigen 1 (NOVA1): Implications in neurological diseases and cancers
Yu Xin, Zheng Li, Heyi Zheng, Jeffery Ho, Matthew T. V. Chan, William K. K. Wu
Cell Proliferation.2017;[Epub] CrossRef - New developments in the pathology of malignant lymphoma. A review of the literature published from January–April 2016
J. Han van Krieken
Journal of Hematopathology.2016; 9(2): 73. CrossRef
 PubReader
PubReader ePub Link
ePub Link-
 Cite this Article
Cite this Article
- Cite this Article
-
- Close
- Download Citation
- Close
- Figure
-
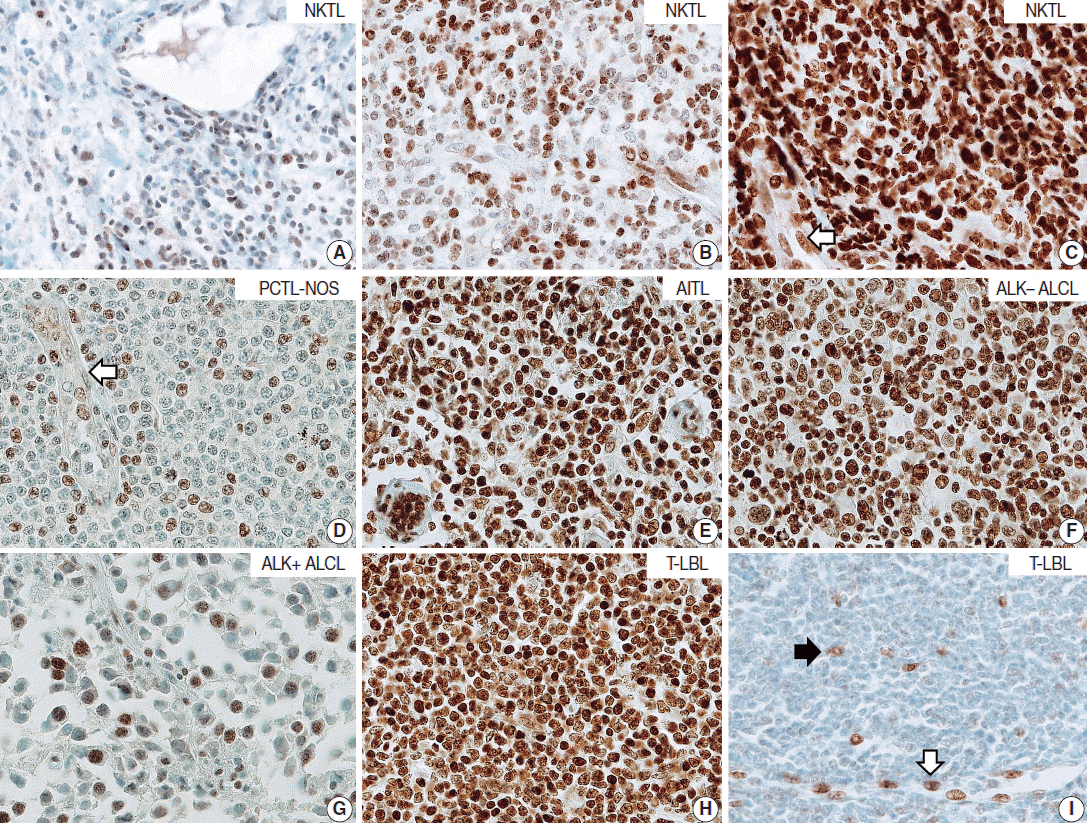
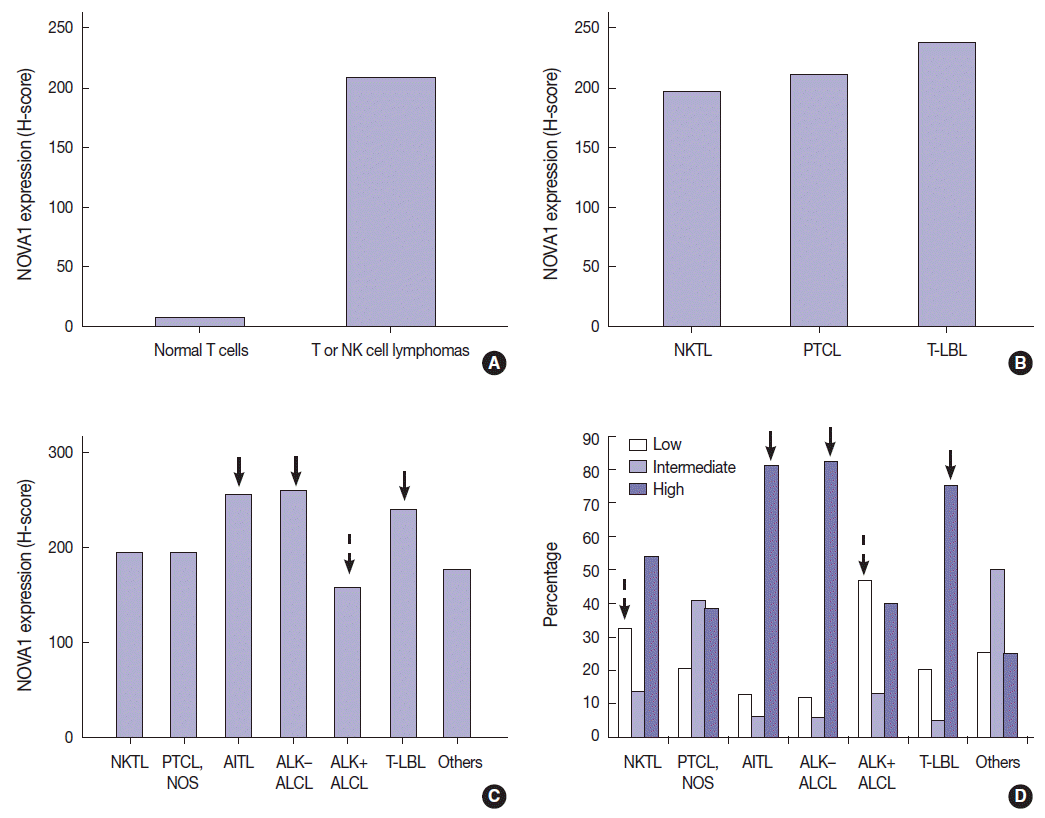
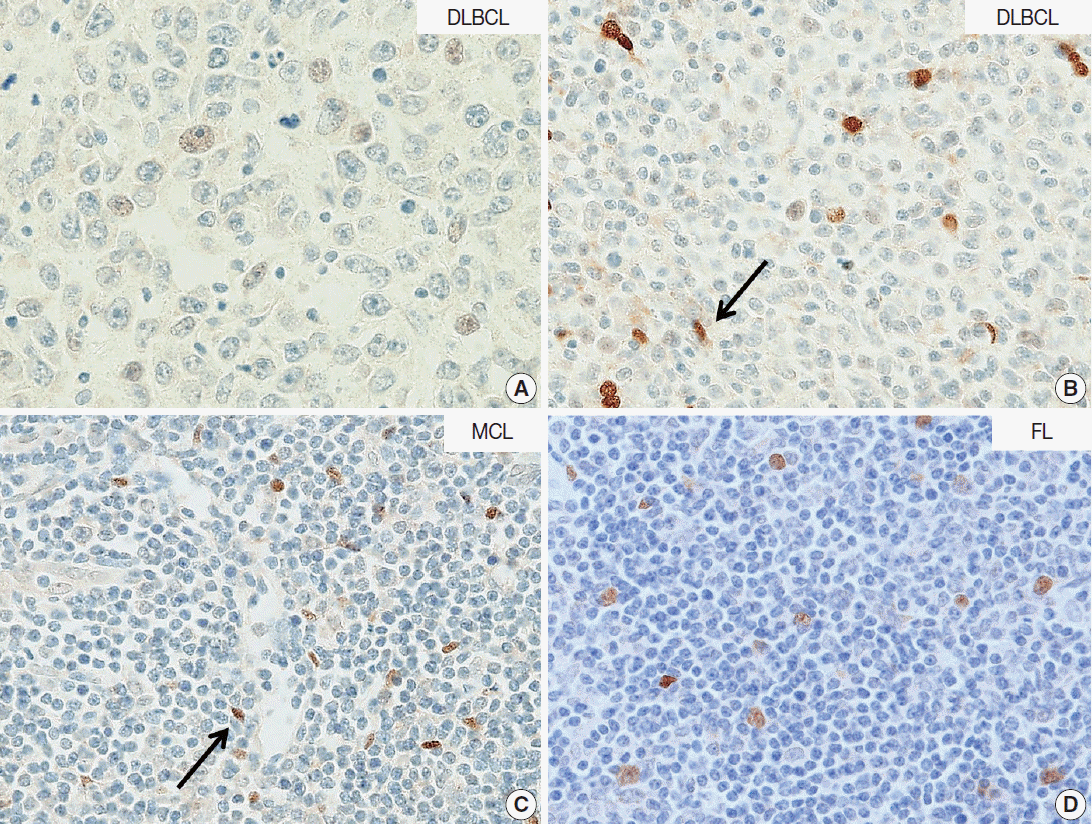
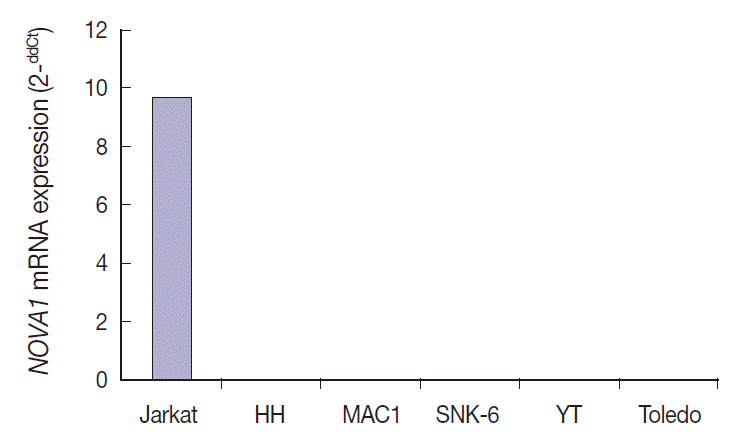
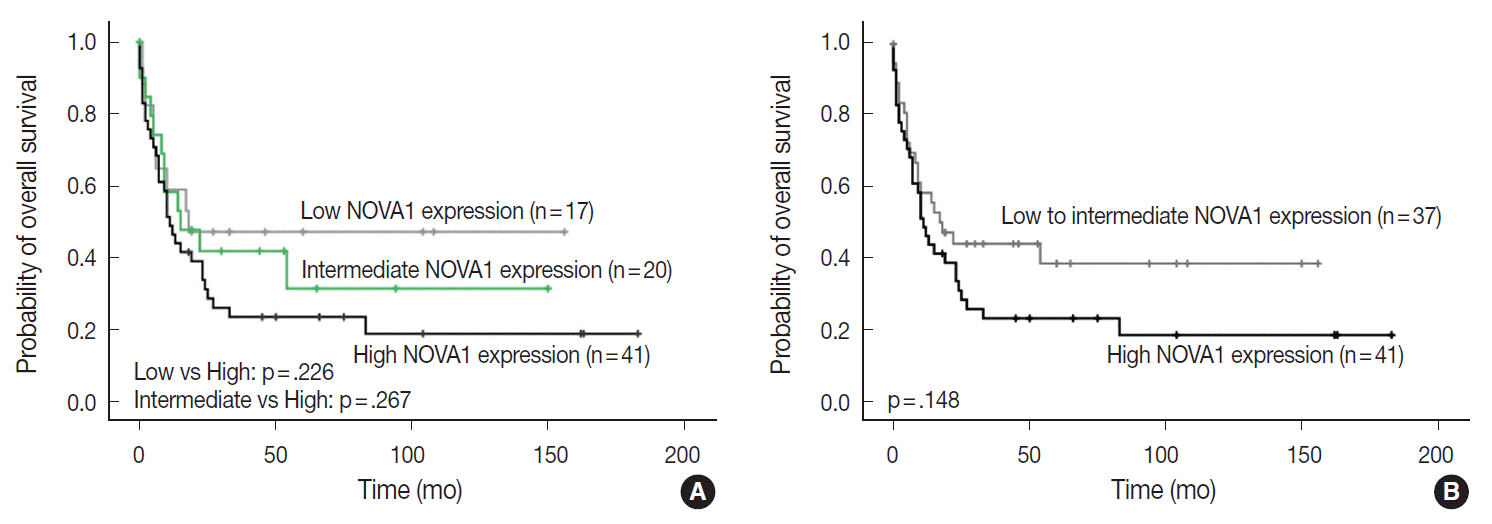





Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
| Tissue category | Case No. | NOVA1 expression |
||||
|---|---|---|---|---|---|---|
| General pattern | Frequency of expression category (%) |
H-score (mean) | ||||
| High | Intermediate | Low | ||||
| Palatine tonsils | 20 | Small numbers of T cells | 0 | 0 | 100 | 7 |
| T- and NK-cell lymphomas | 177 | Strong and diffuse | 56.5 | 18.6 | 24.9 | 209 |
| NKTL | 60 | Lower | 54.2 | 13.6 | 32.2 | 195 |
| PTCL-NOS | 44 | Diverse | 38.6 | 40.9 | 20.5 | 194 |
| AITL | 16 | Higher | 81.2 | 6.2 | 12.5 | 255 |
| ALK-negative ALCL | 17 | Higher | 82.4 | 5.9 | 11.8 | 259 |
| ALK-positive ALCL | 16 | Lower | 40 | 13.3 | 46.7 | 156 |
| T-LBL | 20 | Higher | 75 | 5 | 20 | 237 |
| Other types |
4 | NA | 25 | 50 | 25 | 175 |
| B-Cell lymphomas | 182 | Almost negative | 0 | 0 | 100 | 7 |
| DLBCL | 151 | Almost negative | 0 | 0 | 100 | 8 |
| Other types |
31 | Almost negative | 0 | 0 | 100 | 5 |
| Characteristic | No. (%) (n = 139) |
|---|---|
| Sex (male vs female) | 94 (67.1):45 (32.9) |
| Age (< 60 yr vs ≥ 60 yr) | 91 (65.0):49 (35.0) |
| Subtype | |
| NKTL | 45 (32.4) |
| PTCL | 38 (27.3) |
| AITL | 14 (10.1) |
| ALCL, ALK-negative | 12 (8.6) |
| ALCL, ALK-positive | 12 (8.6) |
| T-LBL | 15 (10.8) |
| Others | 3 (2.2) |
| Primary site of tumor | |
| H&N vs LN vs GI vs soft tissue and bone vs others (solid organs) | 37 (26.4):72 (51.4):9 (6.4):14 (10.0):8 (5.7) |
| Ann Arbor stage (I & II vs III&IV vs not evaluable) | 29 (27.9):77 (55.0):34 (24.3) |
| LDH level (normal vs elevated vs not evaluable) | 39 (27.9):60 (42.9):41 (29.3) |
| BM involvement (absent vs present vs not evaluable) | 77 (55.0):33 (23.6):30 (21.4) |
| IPI score (0–2 vs 3–5 vs not evaluable) | 65 (46.4):43 (30.7):32 (22.9) |
| Initial treatment modality | |
| CTx alone vs CTx with RTx vs RTx alone vs no Tx vs not evaluable | 63 (45.0):27 (19.3):1 (0.7):1 (0.7):48 (34.3) |
| Antibody | Dilution | Type | Company |
|---|---|---|---|
| CD20 | 1:1600 | Mouse | DAKO, Glostrup, Denmark |
| CD3 | 1:200 | Rabbit | LabVision, Fremont, CA, USA |
| CD4 | 1:200 | Rabbit | Cell Marque, Rocklin, CA, USA |
| CD8 | 1:100 | Mouse | Cell Marque, Rocklin, CA, USA |
| CD56 | 1:200 | Rabbit | Cell Marque, Rocklin, CA, USA |
NOVA1, neuro-oncological ventral antigen 1; NK, natural killer; NKTL, extranodal natural killer/T-cell lymphoma, nasal type; PTCL-NOS, peripheral T-cell lymphoma, not otherwise specified; AITL, angioimmunoblastic T-cell lymphoma; ALK, anaplastic lymphoma kinase; ALCL, anaplastic large cell lymphoma; T-LBL, T lymphoblastic leukemia/lymphoma; DLBCL, diffuse large B-cell lymphoma. The T- or NK-cell lymphomas of the “other type” category included hepatosplenic T-cell lymphoma (n = 2), subcutaneous panniculitis-like T-cell lymphoma (n = 1), and cutaneous gamma-delta T-cell lymphoma (n = 1). The case number for this category was too small (only 4 cases), and therefore NOVA1 expression may not be representative for this category; The set of “other type” B-cell lymphomas was composed of follicular lymphoma (n = 14), mantle cell lymphoma (n = 5), marginal zone B-cell lymphoma (n = 7), and Burkitt lymphoma (n = 5).
NK, natural killer; NKTL, extranodal natural killer/T-cell lymphoma, nasal type; PTCL, peripheral T-cell lymphoma; AITL, angioimmunoblastic T-cell lymphoma; ALCL, anaplastic large cell lymphoma; ALK, anaplastic lymphoma kinase; T-LBL, T lymphoblastic leukemia/lymphoma; H&N, head and neck; LN, lymph node; GI, gastrointestinal tract; LDH, lactate dehydrogenase; BM, bone marrow; IPI, International Prognostic Index; CTx, chemotherapy; RTx, radiotherapy; Tx, treatment.

