 E-submission
E-submission
Articles
- Page Path
- HOME > J Pathol Transl Med > Volume 56(4); 2022 > Article
-
Original Article
EGFL7 expression profile in IDH-wildtype glioblastomas is associated with poor patient outcome -
Bruno Henrique Bressan da Costa1,2
 , Aline Paixão Becker3, Luciano Neder1,4, Paola Gyuliane Gonçalves1,5, Cristiane de Oliveira1,5, Allan Dias Polverini6, Carlos Afonso Clara6, Gustavo Ramos Teixeira2,7, Rui Manuel Reis1,8,9, Lucas Tadeu Bidinotto1,2,5
, Aline Paixão Becker3, Luciano Neder1,4, Paola Gyuliane Gonçalves1,5, Cristiane de Oliveira1,5, Allan Dias Polverini6, Carlos Afonso Clara6, Gustavo Ramos Teixeira2,7, Rui Manuel Reis1,8,9, Lucas Tadeu Bidinotto1,2,5 -
Journal of Pathology and Translational Medicine 2022;56(4):205-211.
DOI: https://doi.org/10.4132/jptm.2022.04.22
Published online: June 15, 2022
1Molecular Oncology Research Center, Barretos Cancer Hospital, Barretos, São Paulo, Brazil
2Barretos School of Health Sciences, Dr. Paulo Prata – FACISB, Barretos, São Paulo, Brazil
3The Ohio State University, Department of Radiation Oncology, Columbus, OH, USA
4Department of Pathology, School of Medicine of Ribeirao Preto, University of Sao Paulo, Ribeirão Preto, Sao Paulo, Brazil
5UNESP – Univ. Estadual Paulista, School of Medicine, Department of Pathology, Botucatu, São Paulo, Brazil
6Department of Neurosurgery, Barretos Cancer Hospital, Barretos, São Paulo, Brazil
7Department of Pathology, Barretos Cancer Hospital, Barretos, São Paulo, Brazil
8Life and Health Sciences Research Institute (ICVS), School of Medicine, University of Minho, Braga, Portugal
9ICVS/3B’s – PT Government Associate Laboratory, Braga/Guimarães, Portugal
- Corresponding Author: Lucas Tadeu Bidinotto, PhD, Molecular Oncology Research Center, Barretos Cancer Hospital, Rua Antenor Duarte Villela, 1331, Barretos, São Paulo 14784 400, Brazil, Tel: +55-1733216600 (ext. 7350), Fax: +55-1733216600, E-mail: lucasbidinotto@gmail.com
© 2022 The Korean Society of Pathologists/The Korean Society for Cytopathology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (https://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Background
- Despite the advances in glioblastoma (GBM) treatment, the average life span of patients is 14 months. Therefore, it is urgent to identity biomarkers of prognosis, treatment response, or development of novel treatment strategies. We previously described the association of high epidermal growth factor-like domain multiple 7 (EGFL7) expression and unfavorable outcome of pilocytic astrocytoma patients. The present study aims to analyze the prognostic potential of EGFL7 in GBM isocitrate dehydrogenase (IDH)-wildtype, using immunohistochemistry and in silico approaches.
-
Methods
- Spearman’s correlation analysis of The Cancer Genome Atlas RNA sequencing data was performed. The genes strongly correlated to EGFL7 expression were submitted to enrichment gene ontology and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis. Additionally, EGFL7 expression was associated with patient overall survival. The expression of EGFL7 was analyzed through immunohistochemistry in 74 GBM IDH-wildtype patients’ samples, and was associated with clinicopathological data and overall survival.
-
Results
- In silico analysis found 78 genes strongly correlated to EGFL7 expression. These genes were enriched in 40 biological processes and eight KEGG pathways, including angiogenesis/vasculogenesis, cell adhesion, and phosphoinositide 3-kinase–Akt, Notch, and Rap1 signaling pathways. The immunostaining showed high EGFL7 expression in 39 cases (52.7%). High immunolabelling was significantly associated with low Karnofsky Performance Status and poor overall survival. Cox analysis showed that GBMs IDH-wildtype with high EGFL7 expression presented a higher risk of death compared to low expression (hazard ratio, 1.645; 95% confidence interval, 1.021 to 2.650; p = .041).
-
Conclusions
- This study gives insights regarding the genes that are correlated with EGFL7, as well as biological processes and signaling pathways, which should be further investigated in order to elucidate their role in glioblastoma biology.
- In silico analysis
- We performed in silico analysis in R software, using the TCGA2STAT package [17] and the tools of cBio [18] and DAVID [19] portals.
- In order to evaluate potential pathways related to EGFL7 expression, correlation analysis was performed in cBio portal. RNA sequencing (RNASeq) data of GBM was obtained from The Cancer Genome Atlas (TCGA) (n = 152 patients), and the genes with Spearman’s correlation ≥ |0.6| were considered strongly correlated to EGFL7 expression. Those were submitted to gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis in DAVID portal, and GO/KEGG terms with p ≤ 0.05 were considered as statistically significant.
- Additionally, RSEM-normalized data of expression (RNASeq) of GBM IDH-wildtype of the TCGA was obtained, together with the clinical data (n = 138). The number of reads of all genes were log10-transformed. Further, Z score was calculated for EGFL7, and patients with Z score ≥ 1.0 were classified with high EGFL7 expression. In order to evaluate the prognostic value of EGFL7, overall survival of the patients was analyzed in Kaplan Meier curves using log-rank statistical test.
- Patients
- Glioblastoma IDH-wildtype tissues were obtained from a series of 74 patients who had undergone surgery at Barretos Cancer Hospital. Formalin-fixed, paraffin embedded slides were reviewed by expert neuropathologists (A.P.B., L.N., and G.R.T.), who confirmed glioblastoma IDH-wildtype diagnosis according to 2021 World Health Organization central nervous system classification [1,20] and selected areas for tissue microarray construction. Overall, the age ranged from 26 to 75 years (median of 57 years), who were stratified in 20–59 years old (45 patients) and > 59 years old (29 patients). The average overall survival was 7.61 months (ranging from 0.13 to 45.50 months). All patients were deceased.
- EGFL7 immunohistochemistry
- The EGFL7 immunohistochemistry reactions were performed according to Brunhara et al. (2021) [14]. Briefly, slides with sections of 4 μm were deparaffinized and rehydrated. Then, antigenic retrieval was performed in tris(hydroxymethyl)aminomethane (Tris)–ethylenediaminetetraacetic acid buffer for 20 minutes at 98ºC. Inhibition of endogenous peroxidases and non-specific proteins was performed with the Novolink Polymer Detection System immunohistochemistry kit (Leica Biosystems, Wetzlar, Germany). The rabbit anti-EGFL7 antibody (cat. No. ab115786, Abcam, Cambridge, MA, USA) was applied at a 1:100 dilution on the slide, for 90 minutes at room temperature. Further, immunoreaction product complexes were treated by Novolink Polymer Detection System (30 minutes of post-primary antibody and 30 minutes of polymer at room temperature). Staining was performed using 3,3′-diaminobenzidine and counterstaining was performed with Gill-2 hematoxylin.
- The reactions were analyzed by an expert neuropathologist (L.N.) using intensity (0, negative; 1, weak; 2, medium; and 3, strong) and extension (0, negative; 1, ~25% of mark; 2, between 25% and 50%; and 3, more than 50% of mark) scores. The scores were summed, and the expression was considered low (sum between 0 and 3) or high (sum between 4 and 6) [14]. Endothelial cells were used as positive internal control.
- Statistical analysis
- Immunohistochemistry staining was associated with clinicopathological data (location, age, sex, and Karnofsky Performance Status [KPS]). Finally, overall survival was analyzed using Kaplan Meier curves. Statistical analysis was performed using chi-square tests or Fisher exact test (clinicopathological), log-rank (overall survival), or univariate Cox proportional hazards. Contrasts with a p ≤ .05 were considered statistically significant.
MATERIALS AND METHODS
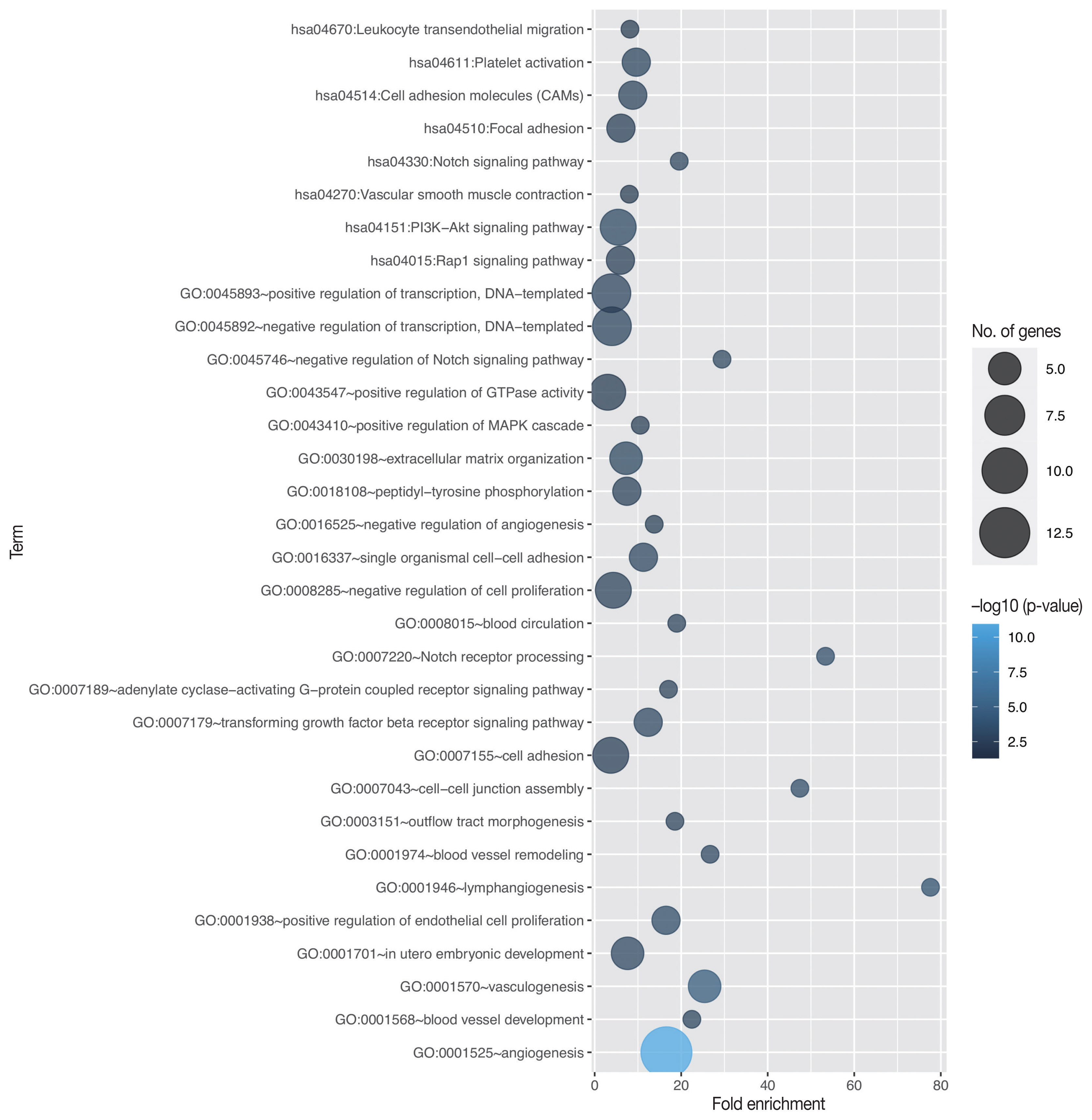
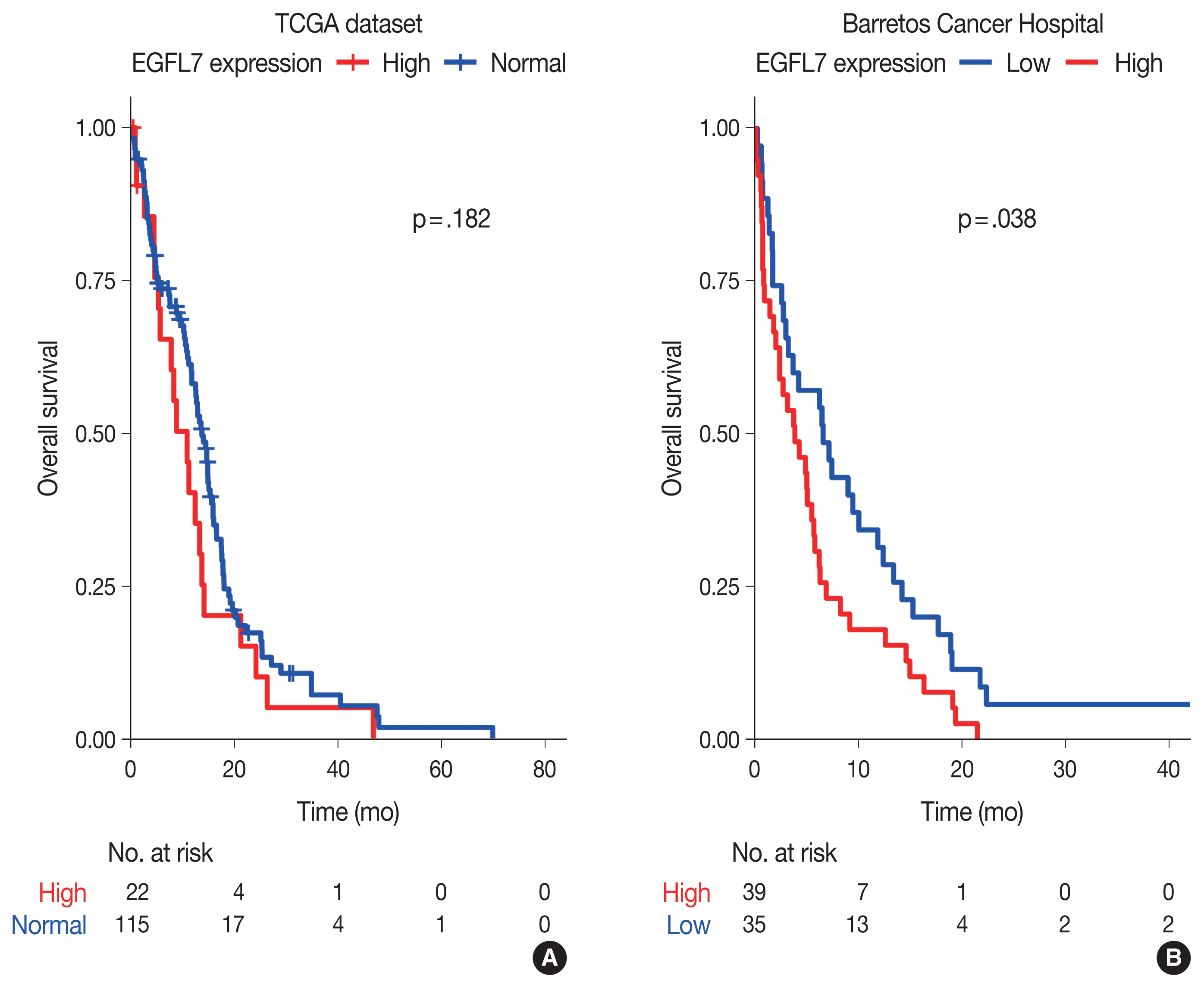
- Through in silico analysis, we found 78 genes significantly associated with EGFL7 expression profile in GBM patients of TCGA database (Supplementary Table S1). These genes were found in 40 enriched GO terms, most of them related to angiogenesis/vasculogenesis, Notch pathway, and cell adhesion (shown in Fig. 1, Supplementary Table S2). Furthermore, pathway analysis using KEGG showed eight enriched terms, mainly related to adhesion, blood cells or blood vessel events, and phosphoinositide 3-kinase (PI3K)–Akt, Notch, and Rap1 signaling pathways (shown in Fig. 1, Supplementary Table S3). Survival analysis of TCGA data showed no difference in overall survival regarding EGFL7 gene expression (p = .182, shown in Fig. 2A).
- We further performed analysis of EGFL7 expression by immunohistochemistry in a series of 74 glioblastomas IDH-wildtype. We found high cytoplasmatic EGFL7 expression in 39 cases (52.7%) (shown in Fig. 3). We further associated EGFL7 immunostaining with patients clinicopathological features (Table 1). High EGFL7 immunolabelling was significantly associated with patients presenting KPS lower or equal to 70 (69.2% of the patients with high EGFL7 expression presented KPS ≤ 70 vs. 45.7% of the patients with low EGFL7 expression, p = .041) (Table 1). EGFL7 expression was not associated with the other clinical features analyzed (sex, location, or age group) (Table 1). Importantly, there was found a poorer overall survival of the patients with high EGFL7 expression (mean of 5.69 months) compared to low EGFL7 expression (mean of 9.95 months, p = 0.038, in log-rank analysis, shown in Fig. 2B). Univariate Cox regression showed that patients with high EGFL7 expression have a 1.645-fold risk of death (95% confidence interval, 1.021 to 2.650; p = .041) compared to patients with low EGFL7 expression.
RESULTS
- In the present study, we analyzed the TCGA database in order to infer genes that may participate in the regulation of EGFL7 expression, as well as evaluated whether EGFL7 expression had a prognostic role in glioblastomas. Our analysis found that patients with high EGFL7 expression generally have low KPS and poor survival. Additionally, our findings give insights of the role and regulation of EGFL7 in this tumor type.
- EGFL7, a downstream target of BMP9-SMAD1/5-mediated signaling, is a secreted angiogenic factor and promotes endothelial expansion through the interference of Notch signaling, ERK activation, and extracellular matrix remodeling [7]. Data of the literature points to the relation of high expression of EGFL7 and poor prognosis in acute myeloid leukemia (AML) [21,22], hepatocellular carcinoma [23], colorectal cancer [10], pancreatic carcinoma [11], and ovarian epithelial cancer [24]. Wang and colleagues additionally found a poor survival of patients with high EGFL7 expression who were EGFRwt [13] mainly by improving the activity of the β-catenin/TCF complex. Li et al. [25] described that EGFL7 expression promotes the proliferation of hepatocellular carcinoma by increasing CKS2. Corroborating these data, EGFR activation (p-EGFR) was found correlated with EGFL7 overexpression in hormone-producing pituitary adenomas [26].
- Our in silico results showed the genes co-expressed with EGFL7 participating mainly in events such as angiogenesis, vasculogenesis, leukocyte transendothelial migration, corroborating the data present in the literature [7,27]. Data also shows enriched genes of Notch pathway (NOTCH4, DLL4, and JAG2). These genes may be subject of further studies to help elucidating the role of this gene in Notch pathway and angiogenesis. In fact, the relation between EGFL7 and Notch pathway remains with conflicting results in the literature. In AML, studies indicate that EGFL7 antagonizes Notch signaling through antagonism to the canonical Notch ligand [28]. On the other hand, silencing of EGFL7 in cutaneous melanoma inactivated Notch pathway, increasing apoptosis and suppressing cell proliferation [29]. Either way, EGFL7 provides its own microenvironment for the migration of endothelial cells, facilitating the budding that leads to angiogenesis [30]. It is known that angiogenesis is one of the hallmarks of cancer, and allows a better supply of blood to the tumor tissue, preventing its cells from having hypoxia/anoxia and/or nutrient deficiency [31].
- Parsatuzumab, a humanized anti-EGFL7 antibody, selectively blocks the interaction between EGFL7 and endothelial cells. There is rationale of using this antibody in the inhibition of vascular growth and, used in combination to anti–vascular endothelial growth factor therapy, inhibition of vascular regrowth after vessel damage, which could reduce tumor vascularization [32]. Given the interesting results of the prognostic role of EGFL7, randomized phase II studies used parsatuzumab in combination with FOLFOX or bevacizumab in the treatment of metastatic colorectal cancer [33] and parsatuzumab in combination with carboplatin, paclitaxel, and bevacizumab for first-line nonsquamous non–small cell lung cancer [34] were performed. The authors found no evidence of the effectiveness of combining parsatuzumab with the proposed treatments, but stress the importance of future studies regarding the mechanisms of EGFL7 and development.
- EGFL7 expression should be considered beyond vascular background since its expression was found in neuronal components in addition to endothelium [35]. Our immunohistochemistry analysis showed that the patients presenting high EGFL7 expression in tumor cells generally have lower KPS and poorer survival. In line, we have previously shown in pilocytic astrocytoma that high EGFL7 expression was associated with unfavorable outcome [14]. Our in silico analysis pointed to the enrichment of PI3K-Akt and Rap1 signaling pathways. In fact, PI3K-Akt pathway has been extensively described in carcinogenesis, including gliomagenesis. Together with mTOR, this pathway regulates several biological processes that are important in growth, metabolism, angiogenesis, autophagy, and chemotherapy resistance in GBM [36]. Importantly, these pathways have been considered for drug design [36]. On the other hand, Rap1 signaling pathway has been less studied in GBM. Rap1 belongs to Ras-related protein family and is involved in cell adhesion and cell junction formation [37] through ERK/mitogen-activated protein kinase signaling and integrin activation [38]. In vitro, Rap1 was activated by integrin through phospholipase D, enhancing cell adhesion and cell proliferation whereas in vivo Rap1 knockdown profoundly altered U373MG glioblastoma growth in xenograft model [39].
- Altogether, our data point to biological functions and pathways related to the increase of tumor vasculature, tumor growth, metabolism, chemotherapy resistance, and changes in cell adhesion and/or cell junction formation. These alterations may be influencing tumor behavior, leading to lower KPS at diagnosis and consequent poor overall survival. However, more molecular studies are warranted in order to elucidate EGFL7 participation in glioblastoma biology.
DISCUSSION
Supplementary Information
Ethics Statement
Written informed consent was obtained from each patient and this study was approved by the ethics committee of Barretos Cancer Hospital (Barretos, Brazil, process CEP 408/2010).
Availability of Data and Material
The datasets generated or analyzed during the current study are available in TCGA repository (https://www.cancer.gov/tcga).
Code Availability
Not applicable.
Conflicts of Interest
The authors declare that they have no potential conflicts of interest.
Author Contributions
Conceptualization: RMR, LTB. Data curation: LTB. Formal analysis: APB, LN, GRT, LTB. Funding acquisition: LTB. Investigation: BHBC, PGG, CO, LTB. Methodology: BHBC, APB, LN, PGG, CO, ADP, CAC. Project administration: RMR, LTB. Resources: RMR, LTB. Supervision: LTB. Validation: RMR, LTB. Visualization: BHBC, APB, LN, GRT, LTB. Writing—original draft: BHBC, LTB. Writing—review & editing: BHBC, PGG, CO, GRT, RMR, LTB.
Funding Statement
This work was supported by The São Paulo Research Foundation (fellowship number 2018/20737-1 to Costa BHB, fellowship number 2017/09749-5 to Gonçalves PG, grant number 2016/21727-4 to Bidinotto LT); Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) fellowship to Oliveira C; CNPq (productivity fellowship to Reis RM and Universal grant number 472447/2013-0 to Bidinotto LT).

- 1. WHO Classification of Tumours Editorial Board Central nervous system tumours. 5th ed. Lyon: International Agency for Research on Cancer, 2021.
- 2. Broniscer A, Gajjar A. Supratentorial high-grade astrocytoma and diffuse brainstem glioma: two challenges for the pediatric oncologist. Oncologist 2004; 9: 197-206. ArticlePubMedPDF
- 3. Hadjipanayis CG, Van Meir EG. Brain cancer propagating cells: biology, genetics and targeted therapies. Trends Mol Med 2009; 15: 519-30. ArticlePubMedPMC
- 4. Paugh BS, Qu C, Jones C, et al. Integrated molecular genetic profiling of pediatric high-grade gliomas reveals key differences with the adult disease. J Clin Oncol 2010; 28: 3061-8. ArticlePubMedPMC
- 5. Ohgaki H. Epidemiology of brain tumors. Methods Mol Biol 2009; 472: 323-42. ArticlePubMed
- 6. Stupp R, Mason WP, van den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med 2005; 352: 987-96. ArticlePubMed
- 7. Richter A, Alexdottir MS, Magnus SH, et al. EGFL7 mediates BMP9-induced sprouting angiogenesis of endothelial cells derived from human embryonic stem cells. Stem Cell Reports 2019; 12: 1250-9. ArticlePubMedPMC
- 8. Parker LH, Schmidt M, Jin SW, et al. The endothelial-cell-derived secreted factor Egfl7 regulates vascular tube formation. Nature 2004; 428: 754-8. ArticlePubMedPDF
- 9. Fitch MJ, Campagnolo L, Kuhnert F, Stuhlmann H. Egfl7, a novel epidermal growth factor-domain gene expressed in endothelial cells. Dev Dyn 2004; 230: 316-24. ArticlePubMedPMC
- 10. Hansen TF, Andersen RF, Olsen DA, Sorensen FB, Jakobsen A. Prognostic importance of circulating epidermal growth factor-like domain 7 in patients with metastatic colorectal cancer treated with chemotherapy and bevacizumab. Sci Rep 2017; 7: 2388.ArticlePubMedPMCPDF
- 11. Shen X, Han Y, Xue X, et al. Epidermal growth factor-like domain 7 promotes cell invasion and angiogenesis in pancreatic carcinoma. Biomed Pharmacother 2016; 77: 167-75. ArticlePubMed
- 12. Huang CH, Li XJ, Zhou YZ, Luo Y, Li C, Yuan XR. Expression and clinical significance of EGFL7 in malignant glioma. J Cancer Res Clin Oncol 2010; 136: 1737-43. ArticlePubMedPDF
- 13. Wang FY, Kang CS, Wang-Gou SY, Huang CH, Feng CY, Li XJ. EGFL7 is an intercellular EGFR signal messenger that plays an oncogenic role in glioma. Cancer Lett 2017; 384: 9-18. ArticlePubMed
- 14. Brunhara BB, Becker AP, Neder L, et al. Evaluation of the prognostic potential of EGFL7 in pilocytic astrocytomas. Neuropathology 2021; 41: 21-8. ArticlePubMedPDF
- 15. Fan C, Yang LY, Wu F, et al. The expression of Egfl7 in human normal tissues and epithelial tumors. Int J Biol Markers 2013; 28: 71-83. ArticlePubMedPDF
- 16. Bidinotto LT, Torrieri R, Mackay A, et al. Copy number profiling of Brazilian astrocytomas. G3 (Bethesda) 2016; 6: 1867-78. ArticlePubMedPMCPDF
- 17. Wan YW, Allen GI, Liu Z. TCGA2STAT: simple TCGA data access for integrated statistical analysis in R. Bioinformatics 2016; 32: 952-4. ArticlePubMedPDF
- 18. Cerami E, Gao J, Dogrusoz U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov 2012; 2: 401-4. ArticlePubMedPMCPDF
- 19. Huang da W, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res 2009; 37: 1-13. ArticlePubMedPMC
- 20. Louis DN, Perry A, Wesseling P, et al. The 2021 WHO classification of tumors of the central nervous system: a summary. Neuro Oncol 2021; 23: 1231-51. ArticlePubMedPMCPDF
- 21. Cheng Z, Dai Y, Pang Y, et al. High EGFL7 expression may predict poor prognosis in acute myeloid leukemia patients undergoing allogeneic hematopoietic stem cell transplantation. Cancer Biol Ther 2019; 20: 1314-8. ArticlePubMedPMC
- 22. Papaioannou D, Shen C, Nicolet D, et al. Prognostic and biological significance of the proangiogenic factor EGFL7 in acute myeloid leukemia. Proc Natl Acad Sci U S A 2017; 114: E4641-7. ArticlePubMedPMC
- 23. Yang C, Wang YL, Sun D, Zhu XL, Li Z, Ni CF. Increased expression of epidermal growth factor-like domain-containing protein 7 is predictive of poor prognosis in patients with hepatocellular carcinoma. J Cancer Res Ther 2018; 14: 867-72. ArticlePubMed
- 24. Oh J, Park SH, Lee TS, Oh HK, Choi JH, Choi YS. High expression of epidermal growth factor-like domain 7 is correlated with poor differentiation and poor prognosis in patients with epithelial ovarian cancer. J Gynecol Oncol 2014; 25: 334-41. ArticlePubMedPMC
- 25. Li Z, Xue TQ, Yang C, Wang YL, Zhu XL, Ni CF. EGFL7 promotes hepatocellular carcinoma cell proliferation and inhibits cell apoptosis through increasing CKS2 expression by activating Wnt/beta-catenin signaling. J Cell Biochem 2018; 119: 10327-37. PubMed
- 26. Liu Q, Zhang J, Gao H, et al. Role of EGFL7/EGFR-signaling pathway in migration and invasion of growth hormone-producing pituitary adenomas. Sci China Life Sci 2018; 61: 893-901. ArticlePubMedPDF
- 27. Nichol D, Shawber C, Fitch MJ, et al. Impaired angiogenesis and altered Notch signaling in mice overexpressing endothelial Egfl7. Blood 2010; 116: 6133-43. ArticlePubMedPMCPDF
- 28. Bill M, Pathmanathan A, Karunasiri M, et al. EGFL7 antagonizes NOTCH signaling and represents a novel therapeutic target in acute myeloid leukemia. Clin Cancer Res 2020; 26: 669-78. ArticlePubMedPDF
- 29. Tang H, Xiao WR, Liao YY, et al. EGFL7 silencing inactivates the Notch signaling pathway; enhancing cell apoptosis and suppressing cell proliferation in human cutaneous melanoma. Neoplasma 2019; 66: 187-96. ArticlePubMed
- 30. Schmidt M, De Maziere A, Smyczek T, et al. The role of Egfl7 in vascular morphogenesis. Novartis Found Symp 2007; 283: 18-28. ArticlePubMedPDF
- 31. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011; 144: 646-74. ArticlePubMed
- 32. Yeung S, Smyczek T, Cheng J, et al. Abstract 3295: inhibiting vascular morphogenesis in tumors: EGFL7 as a novel therapeutic target. Cancer Res 2011; 71: 3295.ArticlePDF
- 33. Garcia-Carbonero R, van Cutsem E, Rivera F, et al. Randomized phase II trial of parsatuzumab (anti-EGFL7) or placebo in combination with FOLFOX and bevacizumab for first-line metastatic colorectal cancer. Oncologist 2017; 22: 375.ArticlePubMedPMCPDF
- 34. von Pawel J, Spigel DR, Ervin T, et al. Randomized phase II trial of parsatuzumab (anti-EGFL7) or placebo in combination with carboplatin, paclitaxel, and bevacizumab for first-line nonsquamous non-small cell lung cancer. Oncologist 2018; 23: 654.ArticlePubMedPMCPDF
- 35. Bicker F, Schmidt MH. EGFL7: a new player in homeostasis of the nervous system. Cell Cycle 2010; 9: 1263-9. ArticlePubMed
- 36. Shahcheraghi SH, Tchokonte-Nana V, Lotfi M, Lotfi M, Ghorbani A, Sadeghnia HR. Wnt/beta-catenin and PI3K/Akt/mTOR signaling pathways in glioblastoma: two main targets for drug design: a review. Curr Pharm Des 2020; 26: 1729-41. ArticlePubMed
- 37. Bos JL, de Rooij J, Reedquist KA. Rap1 signalling: adhering to new models. Nat Rev Mol Cell Biol 2001; 2: 369-77. ArticlePubMedPDF
- 38. Han J, Lim CJ, Watanabe N, et al. Reconstructing and deconstructing agonist-induced activation of integrin alphaIIbbeta3. Curr Biol 2006; 16: 1796-806. PubMed
- 39. Sayyah J, Bartakova A, Nogal N, Quilliam LA, Stupack DG, Brown JH. The Ras-related protein, Rap1A, mediates thrombin-stimulated, integrin-dependent glioblastoma cell proliferation and tumor growth. J Biol Chem 2014; 289: 17689-98. ArticlePubMedPMC
REFERENCES
Figure & Data
References
Citations
- Emerging roles of EGFL family members in neoplastic diseases: Molecular mechanisms and targeted therapies
Xiaoge Gao, Guopeng zhang, Feitong Wang, Wenhui Ruan, Shishuo Sun, Qing Zhang, Xiangye Liu
Biochemical Pharmacology.2025; 236: 116847. CrossRef - Clinicopathological and Prognostic Significance of Epidermal Growth Factor-Like Domain 7 (EGFL7) Overexpression in Primary Central Nervous System Tumor: A Meta-Analysis
IGAP Sasmana, S. Wiranata, IGNAW Kusuma, IGP Supadmanaba, D. M. Wihandani
Innovative Medicine of Kuban.2025; 10(3): 37. CrossRef - Comprehensive Analysis Reveals Epithelial Growth Factor Receptor as a Potential Diagnostic Biomarker in Glioblastoma Multiforme
Amna Makawi, Somia A Khalafallah, Israa M Faris, Mohamed Alfaki
Cureus.2024;[Epub] CrossRef - Role of EGFL7 in human cancers: A review
Cristiane de Oliveira, Paola Gyuliane Gonçalves, Lucas Tadeu Bidinotto
Journal of Cellular Physiology.2023; 238(8): 1756. CrossRef - Low EGFL7 expression is associated with high lymph node spread and invasion of lymphatic vessels in colorectal cancer
Cristiane de Oliveira, Sandra Fátima Fernandes Martins, Paola Gyuliane Gonçalves, Gabriel Augusto Limone, Adhemar Longatto-Filho, Rui Manuel Reis, Lucas Tadeu Bidinotto
Scientific Reports.2023;[Epub] CrossRef
 PubReader
PubReader ePub Link
ePub Link-
 Cite this Article
Cite this Article
- Cite this Article
-
- Close
- Download Citation
- Close
- Figure
-
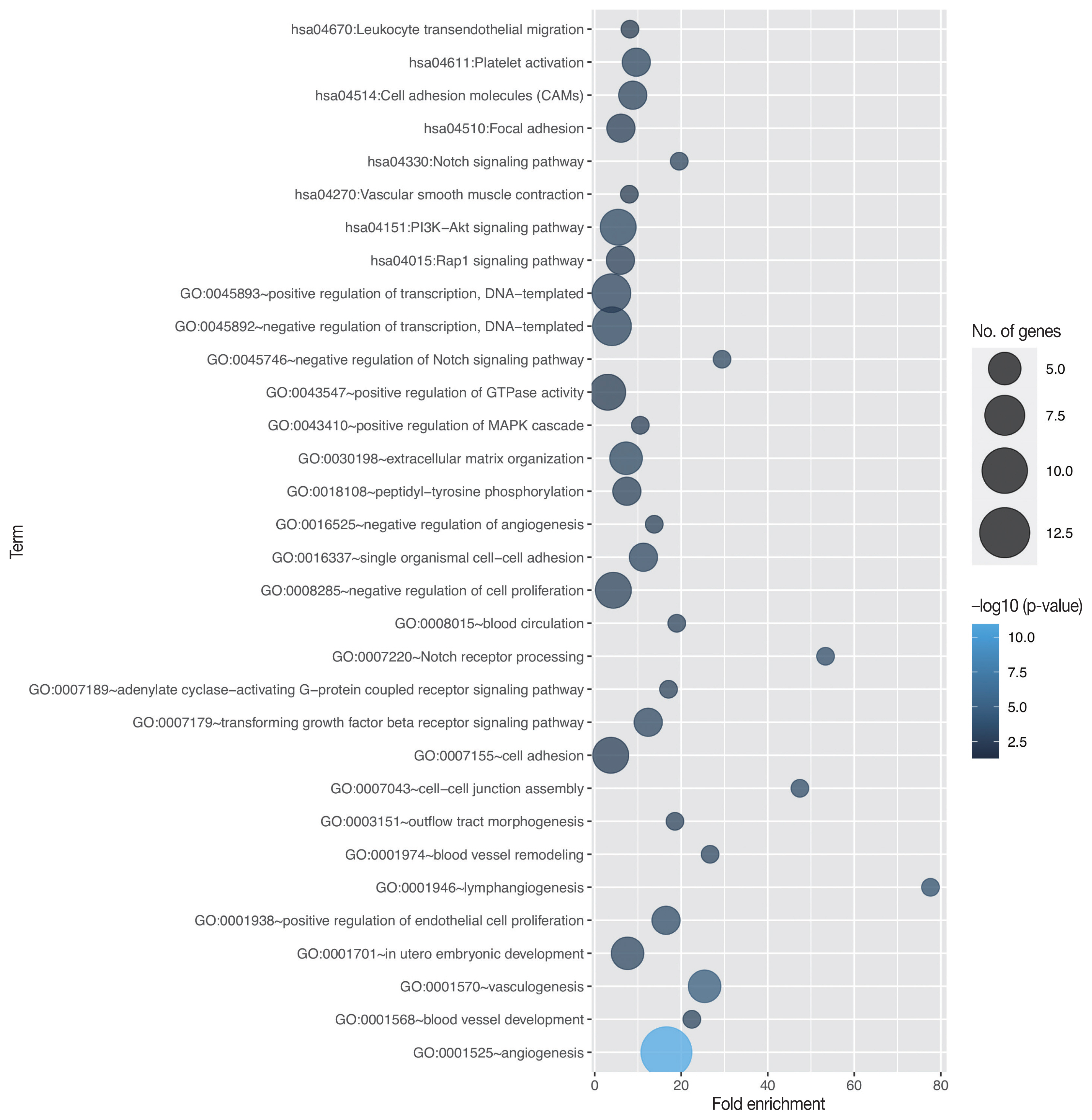
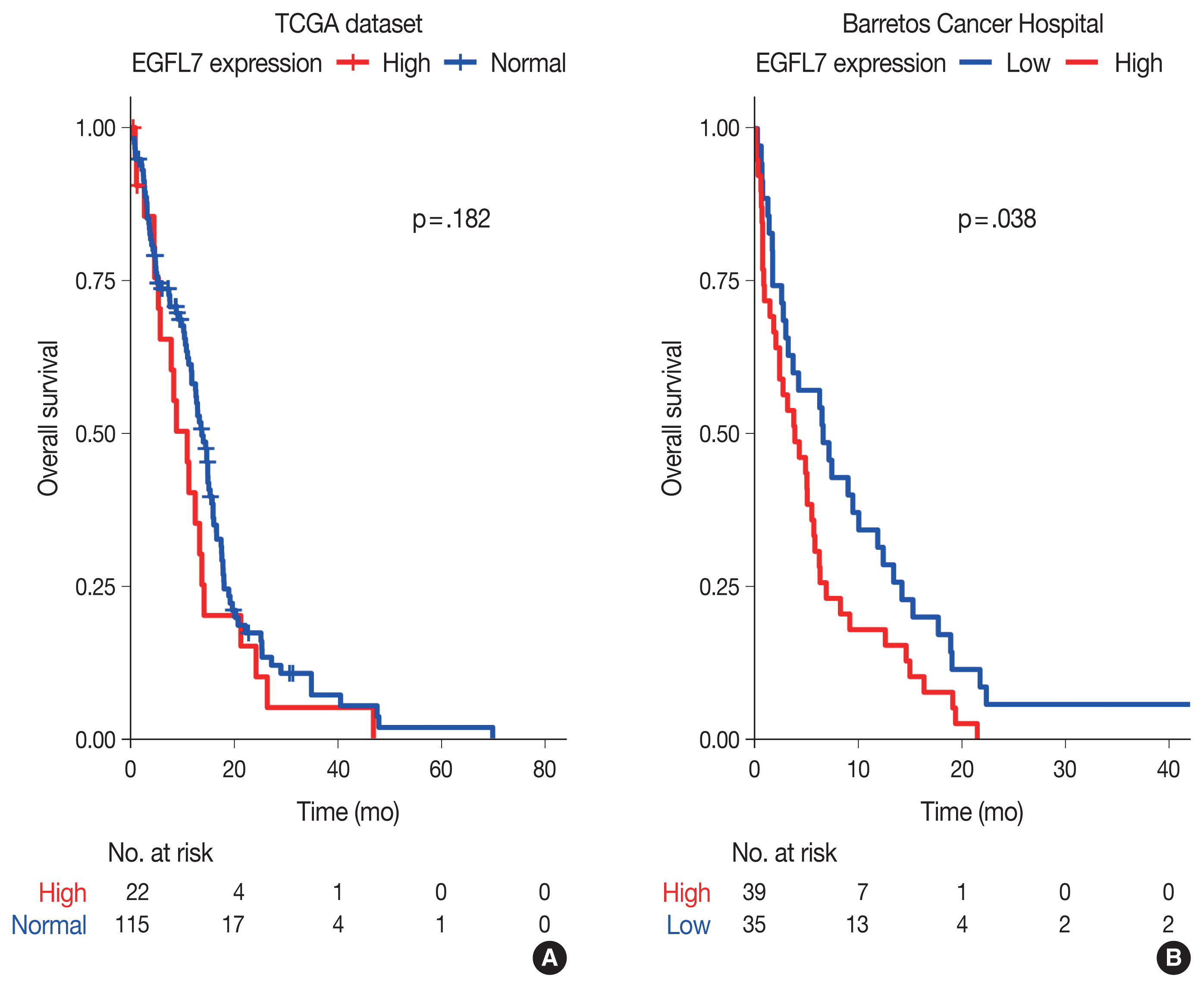
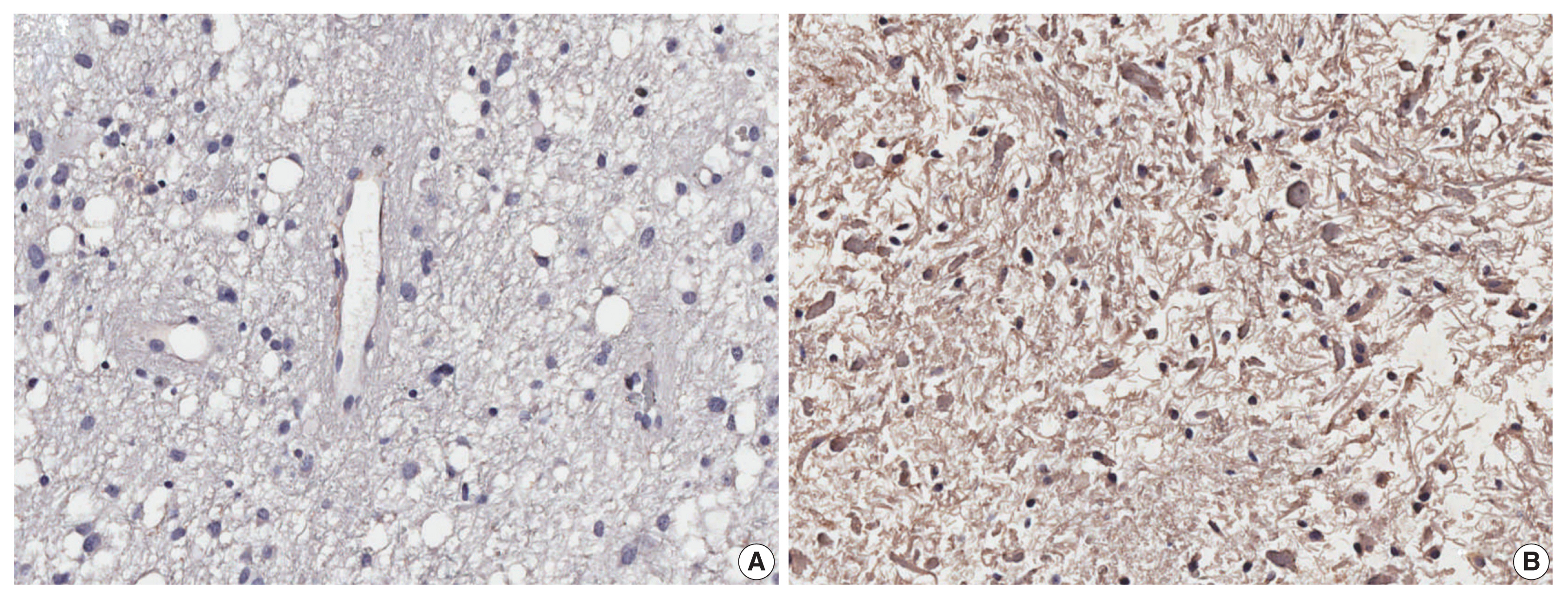



Fig. 1
Fig. 2
Fig. 3
| No. | EGFL7 expression | p-value | ||
|---|---|---|---|---|
|
| ||||
| Low | High | |||
| Sex | ||||
| Female | 29 | 16 (45.7) | 13 (33.3) | 0.276 |
| Male | 45 | 19 (54.3) | 26 (66.7) | |
| Location | ||||
| Frontal | 21 | 12 (34.3) | 9 (23.1) | 0.466 |
| Parietal | 13 | 8 (22.9) | 5 (12.8) | |
| Temporal | 12 | 4 (11.4) | 8 (20.5) | |
| Occipital | 3 | 1 (2.9) | 2 (5.1) | |
| Other | 25 | 10 (28.6) | 15 (38.5) | |
| Age group (yr) | ||||
| 20–59 | 45 | 24 (68.6) | 21 (53.8) | 0.195 |
| > 59 | 29 | 11 (31.4) | 18 (46.2) | |
| KPS | ||||
| ≤ 70 | 43 | 16 (45.7) | 27 (69.2) | 0.041 |
| > 70 | 31 | 19 (54.3) | 12 (30.8) | |
Values are presented as number (%). EGFL7, epidermal growth factor-like domain multiple 7; KPS, Karnofsky Performance Status.

