 E-submission
E-submission
Articles
- Page Path
- HOME > J Pathol Transl Med > Volume 46(1); 2012 > Article
-
Original Article
TNF-α and TNF-β Polymorphisms are Associated with Susceptibility to Osteoarthritis in a Korean Population - Lin Han,, Joo Hyoun Song1,, Jung Hwan Yoon, Yong Gyu Park2, Suk Woo Lee, Yoo Jin Choi, Suk Woo Nam, Jung Young Lee, Won Sang Park
-
Korean Journal of Pathology 2012;46(1):30-37.
DOI: https://doi.org/10.4132/KoreanJPathol.2012.46.1.30
Published online: February 23, 2012
Department of Pathology, The Catholic University of Korea College of Medicine, Seoul, Korea.
1Department of Orthopedics, The Catholic University of Korea College of Medicine, Seoul, Korea.
2Department of Biostatistics, The Catholic University of Korea College of Medicine, Seoul, Korea.
- Corresponding Author: Won Sang Park, M.D. Department of Pathology, The Catholic University of Korea College of Medicine, 505 Banpo-dong, Seocho-gu, Seoul 137-701, Korea. Tel: +82-2-590-1192, Fax: +82-2-537-6586, wonsang@catholic.ac.kr
*Lin Han and Joo Hyoun Song contributed equally to this work.
© 2012 The Korean Society of Pathologists/The Korean Society for Cytopathology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Background
- The tumor necrosis factor (TNF) is believed to play an important role in the pathophysiology of osteoarthritis (OA). Evidence shows that genetic polymorphisms make substantial contributions to the etiology of OA.
-
Methods
- We investigated the genotypes TNF-α and TNF-β in 301 OA patients and 291 healthy subjects as controls. We employed a polymerase chain reaction-restriction fragment length polymorphism and a polymerase chain reaction-single strand conformation polymorphism assay to identify the genotypes TNFA -G308A and TNFB +G252A, respectively.
-
Results
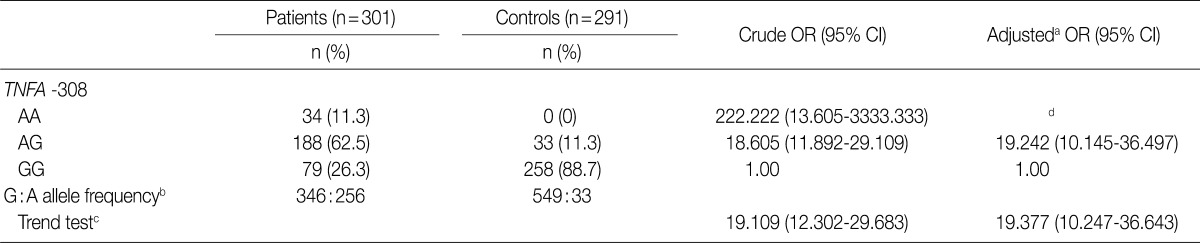
- For TNFA -G308A, the percentages of genotypes GG, AG, and AA were 26.3% (79/301), 62.5% (188/301), and 11.3% (34/301) in OA patients and 88.7% (258/291), 11.3% (33/291), and 0% (0/291) in controls. For TNFB +G252A, the percentages of genotypes GG, AG, and AA were 15.3% (46/301), 41.9% (126/301), and 42.9% (129/301) in OA patients and 12% (35/291), 52.6% (153/291), and 35.4% (103/291) in controls. There were significant differences in genotypes and alleles of TNFA -308 between OA patients and controls (p<0.0001) and in alleles of TNFB +252 (p=0.0325). The risk of OA was significantly higher for carriers of the TNFA -308A allele and the TNFB +252 AA homozygote (p=0.0224).
-
Conclusions
- The results suggest close relationships between TNFA -G308A and TNFB +G252A polymorphisms and individual susceptibility to OA in the Korean population.
- Tissue samples
- We obtained degenerative articular cartilage, meniscus, and ligament tissue specimens from 301 OA patients who received total knee arthroplasty at St. Mary's Hospital of The Catholic University of Korea between 2004 and 2005. All patients were confirmed by radiology and pathology to have OA. Among the 301 patients, 51 (16.9%) were male, and 250 (83.1%) were female. The mean age was 60 at the initial diagnosis. Because these patients received total knee arthroplasty, we obtained the specimens from those patients of Kellgren-Lawrence grade 4 or joint space narrowing grade 4 or higher. We excluded those patients with rheumatoid arthritis (RA), polyarthritis-associated autoimmune diseases, post-traumatic OA, and infection-induced OA. We also excluded those with clinical and radiographic findings suggesting skeletal dysplasia and those with other malignant diseases such as bone tumors, secondary metastasis, alcohol/drug dependence, hepatic failure, and renal failure. The control group consisted of a total of 291 healthy individuals (130 females and 161 males). The mean age was 51. We excluded those individuals with joint pain, a limp, or limited joint movement and those with radiographic signs of joint space narrowing or osteophyte formation. The healthy individuals and patients belonged to the same ethnicity and geographical area. This study was approved by the Institutional Review Board (IRB) of the Catholic University of Korea, College of Medicine (IRB approval No. CUMC10U177).
- DNA extraction
- We extracted DNA templates from the paraffin-embedded knee joint tissue of OA patients. We cut these tissue samples into 4-5 µm slices and dissolved the paraffin with xylene, then washed the xylene with 100% ethanol. We suspended the tissues in an ice-cold Nonidet P-40 lysis buffer solution and treated them with proteinase K. We extracted the DNA by the phenol/chloroform/isoamyl alcohol extraction method and ethanol precipitation, following previous research.14 For healthy individuals, we obtained a leukocyte cell pellet from each blood sample through the Buffy coat by the centrifugation of 2 mL of whole blood. We used the cell pellet for the DNA extraction. We employed the Qiagen DNA blood mini kit (Qiagen, Valencia, CA, USA) and followed the manufacturer's instructions to obtain the genomic DNA. We determined the purity and concentration of the extracted DNA by using the Nanodrop ND-1000 spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA).
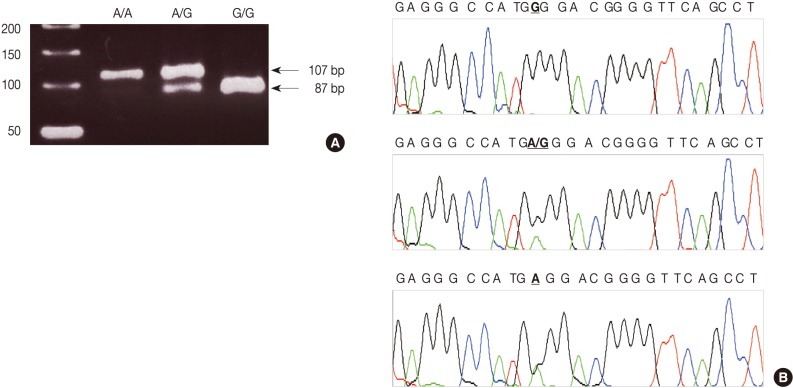
- PCR-RFLP for TNF-α
- We used a PCR-RFLP assay to identify the TNFA -G308A genotype. The primers were sense 5'-AGG CAA TAG GTT TTG AGG GCC AT-3' and antisense 5'-TCC TCC CTG CTC CGA TTC CG-3'. We performed each PCR procedure under standard conditions with a 10 µL PCR mixture containing 1 µL of template DNA, 0.5 µM of each primer, 0.2 µM of each deoxynucleotide triphosphate, 1.5 mM of MgCl2, 0.4 U of the AmpliTaq gold polymerase (Perkin-Elmer, Foster City, CA, USA), and 1 µL of 10× buffer. We denatured the reaction mixture for 12 minutes at 94℃ and incubated it for 40 cycles (denaturation for 30 seconds at 94℃, annealing for 30 seconds at 60℃, and elongation for 30 seconds at 72℃). We continued the final extension for 5 minutes at 72℃. After the amplification, we digested the PCR products with 5 U of the restriction enzyme NCO1 for 4 hours at 37℃. We then separated the digested product on a 3% agarose gel containing ethidium bromide and photographed it by using the Ultra Violet Product Image Store system (Ultraviolet Products, Upland, CA, USA) (Fig. 1A). To ensure the reliability of the RFLP results, we sequenced the PCR products by using the fluorescent dideoxy chain termination method with the ABI 3730XL Analyzer (Applied Biosystems, Foster City, CA, USA) based on the manufacturer's instructions (Fig. 1B).
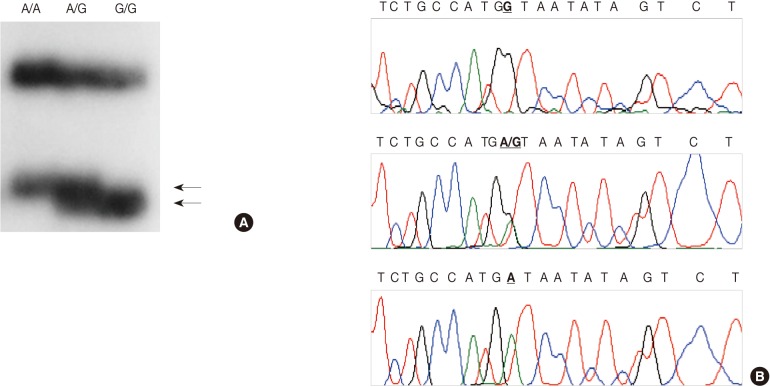
- PCR-SSCP for TNF-β
- We employed a PCR-SSCP assay to identify the TNFB +G252A genotype. The primers were sense 5'-CTC CTG CAC CTG CTG CCT GGA TC-3' and antisense 5'-GAA GAG ACG TTC AGG TGG TGT CAT-3'. We performed each PCR procedure under standard conditions with a 10 µL PCR mixture containing 1 µL of template DNA, 0.5 µM of each primer, 0.2 µM of each deoxynucleotide triphosphate, 1.5 mM of MgCl2, 0.4 U of the AmpliTaq gold polymerase (Perkin-Elmer), 0.5 µCi of [32P]dCTP (Amersham, Buckinghamshire, UK), and 1 µL of 10× buffer. We denatured the reaction mixture for 12 minutes at 95℃ and incubated it for 40 cycles (denaturation for 30 seconds at 95℃, annealing for 30 seconds at 60℃, and elongation for 30 seconds at 72℃). We continued the final extension for 5 minutes at 72℃. After the amplification, we denatured the PCR products for 5 minutes at 95℃ in a 1:1 dilution of a sample buffer solution containing 98% formamide and 5 mmol/L NaOH. We loaded these products onto an SSCP gel (FMC Mutation Detection Enhancement system, Intermountain Scientific, Kaysville, UT, USA) containing 10% glycerol. After the electrophoresis, we transferred the gels to 3 MM Whatman paper and dried them. We then performed the autoradiography by using Kodak X-OMAT film (Eastman Kodak, Rochester, NY, USA) (Fig. 2A). We cut the DNA showing mobility shifts from the dried gels and amplified it for 40 cycles by using the same primer set. We sequenced the PCR products by using the ABI 3730XL Analyzer (Applied Biosystems) (Fig. 2B). Blinded to the status of the study cohort, one of the authors evaluated the results. We randomly selected more than 10% of the sample for repeated assays and found that the results were in complete agreement.
- Statistical analysis
- We conducted a two-tailed Fisher's exact test to determine the differences in the percentages of genotypes and alleles between OA patients and controls. Genotype-specific risk was estimated as an odds ratio at the 95% confidence interval, when we conducted a multiple logistic regression analysis to estimate the interaction between TNF-α and TNF-β. Because both the TNF-α and TNF-β genes are located on chromosome 6p21.3, and the TNF-β polymorphism is associated with TNF-α secretory capacity and the plasma TNF-α level,4,9,10 we analyzed the relationships between the assembled haplotypes and the risk of OA by conducting a logistic regression analysis.
MATERIALS AND METHODS
- TNFA -G308A polymorphism and the risk of OA
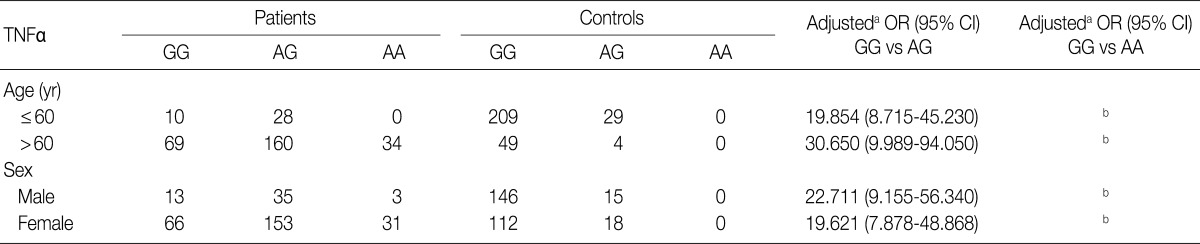
- We considered 301 OA patients and 291 healthy individuals (as controls). Table 1 shows the distributions of genotypes and alleles for the TNFA -G308A polymorphism in OA patients and controls. For healthy individuals, the percentages of genotypes GG, AG, and AA were 88.7% (258/291), 11.3% (33/291), and 0% (0/291), respectively, and the percentages of alleles G and A were 94.3% and 5.7%, respectively. Unexpectedly, healthy individuals did not carry the A/A homozygosity of TNFA -G308A. For OA patients, the percentages of genotypes GG, AG, and AA were 26.3% (79/301), 62.5% (188/301), and 11.3% (34/301), respectively, and the percentages of alleles G and A were 57.5% and 42.5%, respectively. There were significant differences in genotype and allele frequencies for TNFA -308 between OA patients and controls (p<0.0001). We could not estimate the relationships between TNFA -G308A polymorphism genotypes and OA stratified according to age and sex (Table 2).
- TNFB +G252A polymorphism and the risk of OA
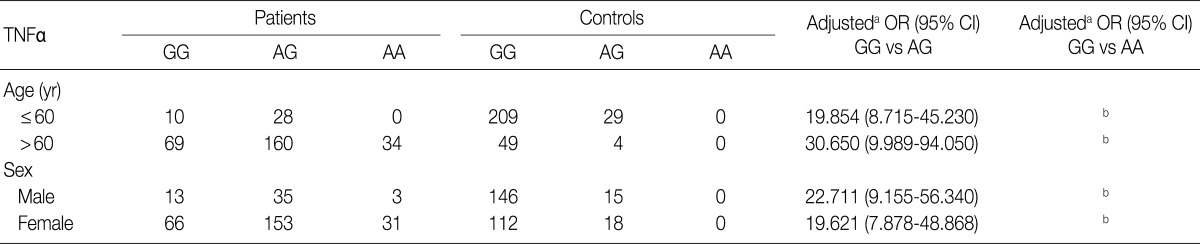
- Table 3 shows the distributions of genotypes and alleles for the TNFB +G252A polymorphism in OA patients and controls. For controls, the percentages of genotypes GG, AG, and AA were 12% (35/291), 52.6% (153/291), and 35.4% (103/291), respectively, and the percentages of alleles G and A were 38.3% and 61.7%, respectively. For OA patients, the percentages of genotypes GG, AG, and AA were 15.3% (46/301), 41.9% (126/301), and 42.9% (129/301), respectively, and the percentages of alleles G and A were 36.2% and 63.8%, respectively. There were significant differences in the percentages of alleles for TNFB +252 between OA patients and controls (p=0.0325). However, there was no significant difference in the genotypic distribution of TNFB +252 (p=0.4542). Table 4 shows the relationship between the TNFB +G252A polymorphism genotype and OA stratified according to age and sex. Because the typical age for OA onset among Koreans is approximately 60, we classified the patients into two age groups: ≤60 as "young" patients and >60 as "old" patients. As shown in Table 4, there was no significant difference between OA patents stratified according to age and sex.
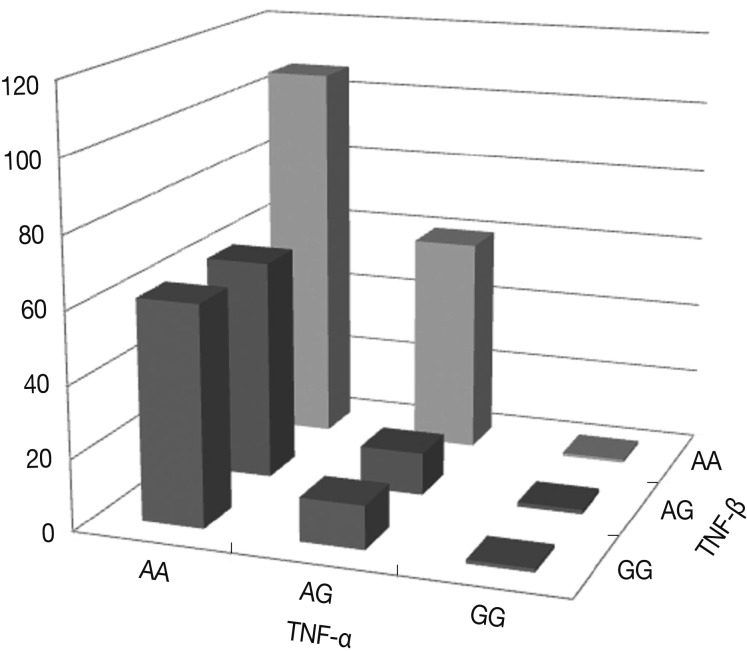
- Haplotypes TNF-α and TNF-β and the risk of OA
- Fig. 3 shows the percentages of genotypes and alleles for the assembled haplotypes TNFA -G308A and TNFB +G252A in OA patients. The results of the logistic regression analysis for the relationships between the assembled haplotypes and the risk of OA indicate that all genotypes with the A allele of the TNF-α gene carried more than a tenfold increase in the risk of OA compared with those genotypes with the G allele of both the TNF-α and TNF-β genes (p<0.0001). There was a significant interaction in terms of the percentages of the assembled haplotypes between TNFA -G308A and TNFB +G252A. In particular, the genotypes with the AA allele of both the TNF-α and TNF-β genes carried more than a hundred fold increase in the risk of OA compared with those genotypes with the GG allele. The risk of OA was significantly higher for carriers of the TNFA -308A allele and the TNFB +252 AA homozygote (p=0.0224).
RESULTS
- Recent studies have shown that some genetic susceptibility contribute to the etiology of OA, including EDG2, ASPN, CALM1, COL2A1, COMP, and FRZB.15 Over the years, it has become evident that the inflammatory cytokine network contributes substantially to the pathogenesis of OA. An increase in the expression of pro-inflammatory cytokines in the patient's cartilage, synovial membrane, and subchondral bone is believed to be linked to the development and progression of structural changes in the OA joint.15 A number of studies have provided evidence of the participation of the TNF family in OA.15 Genetic variations within the TNF promoter may influence the transcription and expression of the TNF. In addition, the up-regulation of TNF gene expression is known to be involved in the pathogenesis of a large variety of illnesses with inflammatory and autoimmune components.
- TNF-α appears to play a pivotal role in the imbalance between anabolic and catabolic processes of OA patients. It can down-regulate the synthesis of major extracellular matrix (ECM) components through the inhibition of the anabolic activity of chondrocytes.16 For instance, TNF-α suppresses the synthesis of link proteins and type II collagen, which are major components of the ECM.17 The TNF also stimulates chondrocytes to release matrix metalloproteinases (MMPs), which have the capacity to degrade cartilage matrix proteins.18 Transgenic mice expressing high concentrations of TNF-α develop arthritis spontaneously.9 SNPs of TNF-α have been found at -863, -857, -308, and -238 sites in the promoter.19 Among the polymorphisms described within the TNF-α promoter, TNFA -G308A has been found to be linked to RA, systemic lupus erythematosus (SLE), and ankylosing spondylitis.13,20 The less common A allele is strongly associated with elevated TNF levels and disease susceptibility in human subjects.21 In this study, the percentages of genotypes at the -G308A site of the TNF-α gene in OA patients were 26.3% for GG, 62.5% for AG, and 11.3% for AA. There was a significant difference in the percentage of the -G308A polymorphism of TNF-α between OA patients and controls (p<0.0001) (Table 1). In addition, there was a significant difference in the allele percentage between OA patients and controls (p<0.0001) (Table 1). These results indicate that the TNFA -308A allele may be a genetic predisposing factor in susceptibility to OA in the Korean population.
- TNF-β is predominantly expressed in B- and T-lymphocytes and natural killer cells. However, its expression has also been described in chondrocytes.22 It has been implicated in lymphoid follicle development and the production of pro-inflammatory cytokines and can facilitate the proliferation of fibroblasts and synoviocytes.23 TNF-β is the homologue of TNF-α, and both bind to TNF-RI and TNF-RII receptors and share many common biological activities. Even at low levels, TNF-β is more likely to induce the secretion of interleukin (IL)-6, IL-8, and MMP3 than TNF-α.24-26 TNF-β is an important pro-inflammatory mediator and biomarker in RA, SLE, and sepsis.24-28 However, the levels of TNF-β production are detected differently in these inflammatory and autoimmune diseases. An important polymorphism influencing TNF-β expression has been found in intron 1 of the gene at the nucleotide position +A252G, and the polymorphism is associated with the risk of RA, SLE, sepsis, pancreatic cancer, and breast cancer.27,28 A high level of the TNFB +252G allele has been found in patients with pancreatic cancer and breast cancer.29 TNFB +252A is associated with the increased risk of developing RA, SLE, and sepsis.24-28 In this study, the percentages of genotypes at the +G252A site of the TNF-β gene in OA patients were 15.3% for GG, 41.9% for AG, and 42.9% for AA. In terms of genotype percentages, there was no significant difference between OA patients and controls (p=0.4542) (Table 3). However, there was a significant difference in the allele percentage at the +G252A site of the TNF-β gene between OA patients and controls (p=0.0325) (Table 3). This suggests that although the A allele of TNFB +A252G is associated with lower TNF-β production,11,12,30 the A allele can increase TNF-α secretory capacity. Thus, the A allele genotype may be a genetic predisposing factor in susceptibility to OA in the Korean population. In addition, there was a significant interaction between TNF-α and TNF-β genes and the occurrence of OA. This suggests that the A allele of the TNF-α gene is the susceptibility gene of OA and that the risk of OA is significantly higher in carriers of the TNFA -308A allele and the TNFB +252 AA homozygote (p=0.0224).
- In conclusion, we demonstrated the relationships of the -G30 8A polymorphism in the TNF-α gene promoter and the +G25 2A polymorphism in intron1 of the TNF-β gene with susceptibility to OA of the knee in a Korean population. Future molecular genetic research should consider larger samples to determine the genetic susceptibility of the TNF-α and TNF-β polymorphisms and identify the mechanisms underlying these polymorphisms.
DISCUSSION
Acknowledgments
Acknowledgments
- 1. Kim I, Kim HA, Seo YI, Song YW, Jeong JY, Kim DH. The prevalence of knee osteoarthritis in elderly community residents in Korea. J Korean Med Sci 2010; 25: 293-298. ArticlePubMedPMCPDF
- 2. Nakajima M, Takahashi A, Kou I, et al. New sequence variants in HLA class II/III region associated with susceptibility to knee osteoarthritis identified by genome-wide association study. PLoS One 2010; 5: e9723.ArticlePubMedPMC
- 3. Shi D, Zheng Q, Chen D, et al. Association of single-nucleotide polymorphisms in HLA class II/III region with knee osteoarthritis. Osteoarthritis Cartilage 2010; 18: 1454-1457. ArticlePubMed
- 4. Carroll MC, Katzman P, Alicot EM, et al. Linkage map of the human major histocompatibility complex including the tumor necrosis factor genes. Proc Natl Acad Sci U S A 1987; 84: 8535-8539. ArticlePubMedPMC
- 5. Körner H, Sedgwick JD. Tumour necrosis factor and lymphotoxin: molecular aspects and role in tissue-specific autoimmunity. Immunol Cell Biol 1996; 74: 465-472. ArticlePubMedPDF
- 6. Abraham LJ, French MA, Dawkins RL. Polymorphic MHC ancestral haplotypes affect the activity of tumour necrosis factor-alpha. Clin Exp Immunol 1993; 92: 14-18. ArticlePubMedPMCPDF
- 7. Fernandes JC, Martel-Pelletier J, Pelletier JP. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002; 39: 237-246. ArticlePubMedPDF
- 8. Abraham LJ, Kroeger KM. Impact of the -308 TNF promoter polymorphism on the transcriptional regulation of the TNF gene: relevance to disease. J Leukoc Biol 1999; 66: 562-566. ArticlePubMedPDF
- 9. Yoon JH, Song JH, Kang YH, et al. TNF-α and TNF-β polymorphisms with susceptibility to gastric cancer in a Korean population. Mol Cell Toxicol 2010; 6: 161-167. ArticlePDF
- 10. Messer G, Spengler U, Jung MC, et al. Polymorphic structure of the tumor necrosis factor (TNF) locus: an NcoI polymorphism in the first intron of the human TNF-beta gene correlates with a variant amino acid in position 26 and a reduced level of TNF-beta production. J Exp Med 1991; 173: 209-219. ArticlePubMedPMCPDF
- 11. Pociot F, Mølvig J, Wogensen L, et al. A tumour necrosis factor beta gene polymorphism in relation to monokine secretion and insulin-dependent diabetes mellitus. Scand J Immunol 1991; 33: 37-49. ArticlePubMed
- 12. Balding J, Kane D, Livingstone W, et al. Cytokine gene polymorphisms: association with psoriatic arthritis susceptibility and severity. Arthritis Rheum 2003; 48: 1408-1413. ArticlePubMed
- 13. Lee JY, Dong SM, Kim SY, Yoo NJ, Lee SH, Park WS. A simple, precise and economical microdissection technique for analysis of genomic DNA from archival tissue sections. Virchows Arch 1998; 433: 305-309. ArticlePubMedPDF
- 14. Ikegawa S. New gene associations in osteoarthritis: what do they provide, and where are we going? Curr Opin Rheumatol 2007; 19: 429-434. ArticlePubMed
- 15. Kapoor M, Martel-Pelletier J, Lajeunesse D, Pelletier JP, Fahmi H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat Rev Rheumatol 2011; 7: 33-42. ArticlePubMedPDF
- 16. Séguin CA, Bernier SM. TNFalpha suppresses link protein and type II collagen expression in chondrocytes: role of MEK1/2 and NF-kappaB signaling pathways. J Cell Physiol 2003; 197: 356-369. PubMed
- 17. Lefebvre V, Peeters-Joris C, Vaes G. Modulation by interleukin 1 and tumor necrosis factor alpha of production of collagenase, tissue inhibitor of metalloproteinases and collagen types in differentiated and dedifferentiated articular chondrocytes. Biochim Biophys Acta 1990; 1052: 366-378. PubMed
- 18. Keffer J, Probert L, Cazlaris H, et al. Transgenic mice expressing human tumour necrosis factor: a predictive genetic model of arthritis. EMBO J 1991; 10: 4025-4031. ArticlePubMedPMCPDF
- 19. Jiménez-Morales S, Velázquez-Cruz R, Ramírez-Bello J, et al. Tumor necrosis factor-alpha is a common genetic risk factor for asthma, juvenile rheumatoid arthritis, and systemic lupus erythematosus in a Mexican pediatric population. Hum Immunol 2009; 70: 251-256. PubMed
- 20. Kroeger KM, Carville KS, Abraham LJ. The -308 tumor necrosis factor-alpha promoter polymorphism effects transcription. Mol Immunol 1997; 34: 391-399. PubMed
- 21. Shlopov BV, Gumanovskaya ML, Hasty KA. Autocrine regulation of collagenase 3 (matrix metalloproteinase 13) during osteoarthritis. Arthritis Rheum 2000; 43: 195-205. ArticlePubMed
- 22. O'Rourke KP, O'Donoghue G, Adams C, et al. High levels of Lymphotoxin-Beta (LT-Beta) gene expression in rheumatoid arthritis synovium: clinical and cytokine correlations. Rheumatol Int 2008; 28: 979-986. ArticlePubMedPDF
- 23. Calmon-Hamaty F, Combe B, Hahne M, Morel J. Lymphotoxin alpha stimulates proliferation and pro-inflammatory cytokine secretion of rheumatoid arthritis synovial fibroblasts. Cytokine 2011; 53: 207-214. ArticlePubMed
- 24. Santos MJ, Fernandes D, Caetano-Lopes J, et al. Lymphotoxin-alpha 252 A>G polymorphism: a link between disease susceptibility and dyslipidemia in rheumatoid arthritis? J Rheumatol 2011; 38: 1244-1249. ArticlePubMed
- 25. Kim HY, Lee SH, Yang HI, et al. TNFB gene polymorphism in patients with systemic lupus erythematosus in Korean. Korean J Intern Med 1995; 10: 130-136. ArticlePubMedPMC
- 26. Temple SE, Almeida CM, Cheong KY, Wunderink RG, Waterer GW. A diplotype in the lymphotoxin alpha gene is associated with differential expression of LTA mRNA induced by Gram-positive and Gram-negative bacteria. Int J Immunogenet 2007; 34: 157-160. ArticlePubMed
- 27. Barber MD, Powell JJ, Lynch SF, Gough NJ, Fearon KC, Ross JA. Two polymorphisms of the tumour necrosis factor gene do not influence survival in pancreatic cancer. Clin Exp Immunol 1999; 117: 425-429. ArticlePubMedPMCPDF
- 28. Kohaar I, Tiwari P, Kumar R, et al. Association of single nucleotide polymorphisms (SNPs) in TNF-LTA locus with breast cancer risk in Indian population. Breast Cancer Res Treat 2009; 114: 347-355. ArticlePubMedPDF
- 29. Temple SE, Cheong KY, Almeida CM, Price P, Waterer GW. Polymorphisms in lymphotoxin alpha and CD14 genes influence TNFalpha production induced by Gram-positive and Gram-negative bacteria. Genes Immun 2003; 4: 283-288. ArticlePubMedPDF
- 30. Pociot F, Briant L, Jongeneel CV, et al. Association of tumor necrosis factor (TNF) and class II major histocompatibility complex alleles with the secretion of TNF-alpha and TNF-beta by human mononuclear cells: a possible link to insulin-dependent diabetes mellitus. Eur J Immunol 1993; 23: 224-231. ArticlePubMed
REFERENCES
TNF, tumor necrosis factor; OA, osteoarthritis; OR, odds ratio; CI, confidence interval.
aAdjusted for age (yr) and sex; bTwo-sided Fisher's exact test: for allele frequencies, p<0.0001; for genotype distribution, p<0.0001; cCalculated in the logistic regression model using the number of A alleles in the genotypes as a continuous variable; dCannot be estimated due to zero frequency in control group.

TNF, tumor necrosis factor; OA, osteoarthritis; OR, odds ratio; CI, confidence interval
aAdjusted for age (yr) and sex; bTwo-sided Fisher's exact test: for allele frequencies, p=0.0325; for genotype distribution, p=0.4542; cCalculated in the logistic regression model using the number of A alleles in the genotypes as a continuous variable.
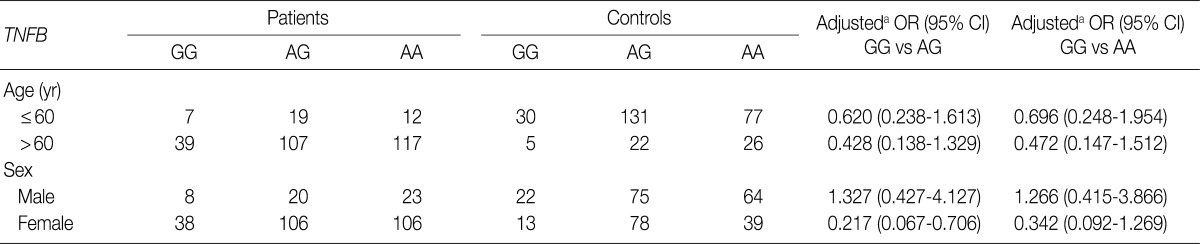
Figure & Data
References
Citations
- Chondroprotective effects of madecassoside incorporated alginate di-aldehyde gelatin hydrogel on IL-1 β induced in vitro osteoarthritic model
Jalaja Aswathy, Rajalekshmi Resmi, Josna Joseph, Roy Joseph, Annie John, Sabulal Baby, Annie Abraham
International Journal of Biological Macromolecules.2026; 335: 149228. CrossRef - Osteoarthritis and analgesic consumption in haemochromatosis HFE C282Y homozygotes with normal or low iron parameters
Jens Helby, Mathis Mottelson, Stig Egil Bojesen, Børge Grønne Nordestgaard, Mikkel Faurschou, Jesper Petersen, Christina Ellervik, Marie Warny, Andreas Glenthøj
Nature Communications.2026;[Epub] CrossRef - Causal effects of circulating inflammatory proteins on knee and hip osteoarthritis: A two sample Mendelian randomization study
Rongjie Lin, Qingcong Zheng, Rui Zhang, Quan Liu, Jianlong Lin, Shaohuang Weng, Min Chen
Clinical Rheumatology.2025; 44(1): 465. CrossRef - Association of Dopamine D2 Receptor (DRD2) C939T and a Lack of Association of Tumor Necrosis Factor-β (TNF-β) +A252G Polymorphisms with Susceptibility to Migraine in A Northern Iranian Population
Negar Gorjizadeh, Payam Saadat, Abbas Azadmehr, Saeed Irian
OBM Genetics.2025; 09(03): 1. CrossRef - The Exosomes of Stem Cells from Human Exfoliated Deciduous Teeth Suppress Inflammation in Osteoarthritis
Chuang-Yu Lin, Parichart Naruphontjirakul, Te-Yang Huang, Yi-Chia Wu, Wei-Hsuan Cheng, Wen-Ta Su
International Journal of Molecular Sciences.2024; 25(16): 8560. CrossRef - Impacts of mesenchymal stem cells and hyaluronic acid on inflammatory indicators and antioxidant defense in experimental ankle osteoarthritis
Usama Ismaeil Hagag, Fatma Mohamed Halfaya, Hessah Mohammed Al-Muzafar, Suhailah Saud Al-Jameel, Kamal Adel Amin, Wael Abou El-Kheir, Emad A Mahdi, Gamal Abdel-Nasser Ragab Hassan, Osama Mohamed Ahmed
World Journal of Orthopedics.2024; 15(11): 1056. CrossRef - Health benefits of astaxanthin against age-related diseases of multiple organs: A comprehensive review
Phaniendra Alugoju, V. K. D. Krishna Swamy, Naga Venkata Anusha Anthikapalli, Tewin Tencomnao
Critical Reviews in Food Science and Nutrition.2023; 63(31): 10709. CrossRef - Complete sequence and comparative analysis of the mitochondrial genome of the rare and endangered Clematis acerifolia, the first clematis mitogenome to provide new insights into the phylogenetic evolutionary status of the genus
Dan Liu, Kai Qu, Yangchen Yuan, Zhiheng Zhao, Ying Chen, Biao Han, Wei Li, Yousry A. El-Kassaby, Yangyang Yin, Xiaoman Xie, Boqiang Tong, Hongshan Liu
Frontiers in Genetics.2023;[Epub] CrossRef - Intra-articular Treatment with Triamcinolone Hexacetonide Associated with Gold Nanoparticles Reduces Cartilage Degeneration in an Animal Model of Osteoarthritis
Daniela Pacheco dos Santos Haupenthal, Marcus Barg Resmini, Leandro Almeida Da Silva, Mateus Cardoso Colares, Laura de Roch Casagrande, Ligia Milanez Venturini, Thiago Antônio Moretti de Andrade, Fernando Russo Costa do Bomfim, Anand Thirupathi, Paulo Emi
Current Drug Targets.2023; 24(3): 287. CrossRef - Vitamin D Receptor Gene Polymorphisms and Risk of Knee Osteoarthritis: Possible Correlations with TNF-α, Macrophage Migration Inhibitory Factor, and 25-Hydroxycholecalciferol Status
Mohammed H. Hassan, Amer Alkot Mostafa Elsadek, Marwa Ahmed Mahmoud, Bakheet E. M. Elsadek
Biochemical Genetics.2022; 60(2): 611. CrossRef - Complement System Inhibition Modulates the Inflammation Induced by the Venom of Premolis semirufa, an Amazon Rainforest Moth Caterpillar
Joel J. M. Gabrili, Isadora Maria Villas-Boas, Giselle Pidde, Carla Cristina Squaiella-Baptistão, Trent M. Woodruff, Denise V. Tambourgi
International Journal of Molecular Sciences.2022; 23(21): 13333. CrossRef - Azilsartan prevented AGE‐induced inflammatory response and degradation of aggrecan in human chondrocytes through inhibition of Sox4
Jie Lei, Mengyin He, Liangzhou Xu, Chengjian He, Jie Li, Wei Wang
Journal of Biochemical and Molecular Toxicology.2021;[Epub] CrossRef - TNF-α Carried by Plasma Extracellular Vesicles Predicts Knee Osteoarthritis Progression
Xin Zhang, Ming-Feng Hsueh, Janet L. Huebner, Virginia B. Kraus
Frontiers in Immunology.2021;[Epub] CrossRef - Genetic association between TNF-α G-308A and osteoarthritis in Asians: A case–control study and meta-analysis
Chih-Chien Wang, Chih-Yun Huang, Meng-Chang Lee, Dung-Jang Tsai, Chia-Chun Wu, Sui-Lung Su, Mingqing Xu
PLOS ONE.2021; 16(11): e0259561. CrossRef - Sex-, Age-, and Metabolic Disorder-Dependent Distributions of Selected Inflammatory Biomarkers among Community-Dwelling Adults
So Mi Jemma Cho, Hokyou Lee, Jee-Seon Shim, Hyeon Chang Kim
Diabetes & Metabolism Journal.2020; 44(5): 711. CrossRef - Tumor necrosis factor-α: Molecular assessment of gene expression, genetic variants and serum level in Egyptian patients with knee osteoarthritis
Nermin Raafat, Amal F. Gharib, Doaa S. Atta, Shimaa M. AbdElwahab, Doaa M. Sharaf
Gene Reports.2020; 21: 100922. CrossRef - Effect of a functional variant of tumor necrosis factor‐β gene in temporomandibular disorders: A pilot study
Kaan Yerliyurt, Ayse Feyda Nursal, Akin Tekcan, Nevin Karakus, Mehmet K. Tumer, Serbulent Yigit
Journal of Clinical Laboratory Analysis.2019;[Epub] CrossRef - Interleukin-6 (-572) and Tumor Necrosis Factor α (-308 and -238) Polymorphisms and Risk of Knee Osteoarthritis in a Pakistani Population: A Case–Control Study
Syeda Kanwal Batool Naqvi, Maria Bibi, Iram Murtaza, Qamar Javed
Iranian Journal of Science and Technology, Transactions A: Science.2019; 43(4): 1485. CrossRef - Role of resistin genetic variations in knee osteoarthritis pathogenesis, a cross sectional study
Syeda Kanwal Batool Naqvi, Iram Murtaza, Qamar Javed
Molecular Biology Reports.2019; 46(3): 2657. CrossRef -
Relationship Between Genetic Polymorphisms of the
TNF
Gene and Hallux Valgus Susceptibility
Jie Yang, Junhu Wang, Xiaojun Liang, Hongmou Zhao, Jun Lu, Qiang Ma, Feng Tian
Genetic Testing and Molecular Biomarkers.2019; 23(6): 380. CrossRef - Functional status and severity of osteoarthritis in elderly is associated to the polymorphism of TNFA gene
Marcos Tadeu Parron Fernandes, Karen Barros Parron Fernandes, Fernanda Freitas Anibal, Walquíria Shimoya-Bittencourt, Viviane Martins Santos, Priscila Daniele de Oliveira Perrucini, Regina Célia Poli-Frederico
Advances in Rheumatology.2019;[Epub] CrossRef - Association of TNF-α-308 G > A and −238G > A polymorphisms with knee osteoarthritis risk: A case-control study and meta-analysis
Mohammad Reza Sobhan, Masoud Mahdinezhad-Yazdi, Kazem Aghili, Masoud Zare-Shehneh, Shohreh Rastegar, Jalal Sadeghizadeh-Yazdi, Hossein Neamatzadeh
Journal of Orthopaedics.2018; 15(3): 747. CrossRef - Association between tumor necrosis factor alpha rs1800629 polymorphism and risk of osteoarthritis in a Chinese population
Jie Chen, Yu Wu, Jiannong Yu, Jinming Shen
Brazilian Journal of Medical and Biological Research.2018;[Epub] CrossRef - Association of cytokine gene polymorphisms with osteoarthritis susceptibility
Otilia Rogoveanu, Daniela Calina, Mihai Cucu, Florin Burada, Anca Docea, Simona Sosoi, Emilian Stefan, Mihai Ioana, Emilia Burada
Experimental and Therapeutic Medicine.2018;[Epub] CrossRef - TNF‑α increases the expression of inflammatory factors in synovial fibroblasts by inhibiting the PI3K/AKT pathway in a rat model of monosodium iodoacetate‑induced osteoarthritis
Hongxi Li, Shujuan Xie, Yunlong Qi, Huazhe Li, Rui Zhang, Yongyun Lian
Experimental and Therapeutic Medicine.2018;[Epub] CrossRef - The single-nucleotide polymorphism (SNP) of tumor necrosis factor α −308G/A gene is associated with early-onset primary knee osteoarthritis in an Egyptian female population
Sahar M. Abdel Galil, Nillie Ezzeldin, Faten Fawzy, Mohamed El-Boshy
Clinical Rheumatology.2017; 36(11): 2525. CrossRef - Roxatidine Attenuates Degradation of Extracellular Matrix
Renhao Ze, Shangyu Wang, Mao Xie, Bo Zhang, Xin Tang, Jin Li
Biomedicine & Pharmacotherapy.2017; 95: 1156. CrossRef - The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis
Piotr Wojdasiewicz, Łukasz A. Poniatowski, Dariusz Szukiewicz
Mediators of Inflammation.2014; 2014: 1. CrossRef - Meta-analysis of tumor necrosis factor alpha -308 polymorphism and knee osteoarthritis risk
Suotang Kou, Yaochi Wu
BMC Musculoskeletal Disorders.2014;[Epub] CrossRef - Astaxanthin reduces matrix metalloproteinase expression in human chondrocytes
Wei-Ping Chen, Yan Xiong, Yong-Xiang Shi, Peng-Fei Hu, Jia-Peng Bao, Li-Dong Wu
International Immunopharmacology.2014; 19(1): 174. CrossRef - Variations in the TNFα gene and their interactions with the IL4R and IL10 genes in relation to hand osteoarthritis
Satu Hämäläinen, Svetlana Solovieva, Tapio Vehmas, Päivi Leino-Arjas, Ari Hirvonen
BMC Musculoskeletal Disorders.2014;[Epub] CrossRef - Tumor necrosis factor gene polymorphisms and endometriosis in Asians: a systematic review and meta-analysis
Jiangtao Lyu, Hua Yang, Jinghe Lang, Xianjie Tan
Chinese Medical Journal.2014; 127(9): 1761. CrossRef - Association analysis of two candidate polymorphisms in the Tumour Necrosis Factor-α gene with osteoarthritis in a Chinese population
Bin Ji, Jixiang Shi, Xiangyu Cheng, Junjie Zhou, Qiang Zhou, Chengfu Cao, Jinhui Pang
International Orthopaedics.2013; 37(10): 2061. CrossRef
 PubReader
PubReader Cite this Article
Cite this Article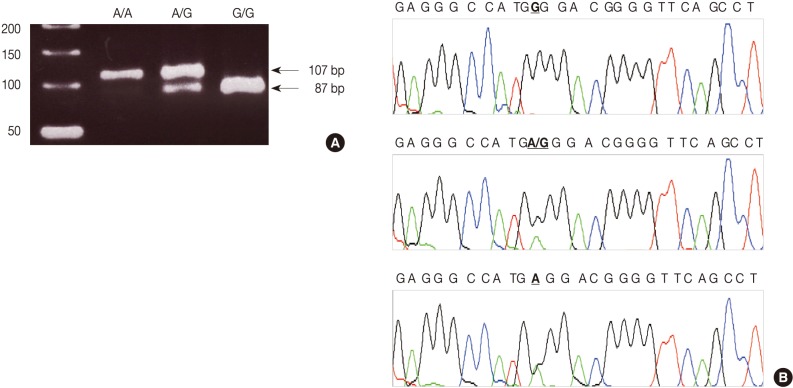
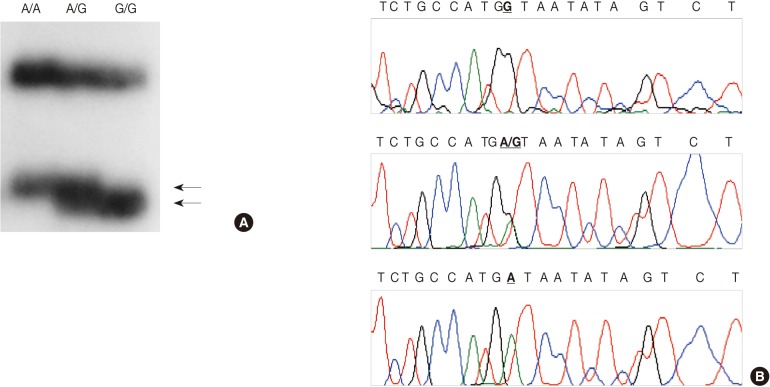
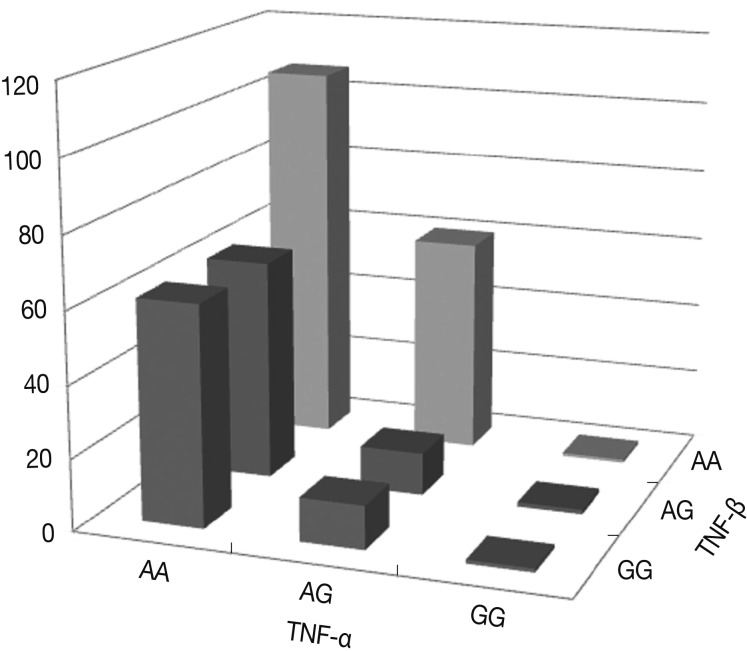



Fig. 1
Fig. 2
Fig. 3



aAdjusted for age (yr) and sex; bTwo-sided Fisher's exact test: for allele frequencies, p<0.0001; for genotype distribution, p<0.0001; cCalculated in the logistic regression model using the number of A alleles in the genotypes as a continuous variable; dCannot be estimated due to zero frequency in control group.
aAdjusted for the other covariates (age [yr] as a continuous variable) presented in this table in a logistic regression model for each stratum; bCannot be estimated due to zero frequency in control group.
aAdjusted for age (yr) and sex; bTwo-sided Fisher's exact test: for allele frequencies, p=0.0325; for genotype distribution, p=0.4542; cCalculated in the logistic regression model using the number of A alleles in the genotypes as a continuous variable.
aAdjusted for the other covariates (age [yr] as a continuous variable) presented in this table in a logistic regression model for each stratum.

