 E-submission
E-submission
Articles
- Page Path
- HOME > J Pathol Transl Med > Volume 50(5); 2016 > Article
-
Original Article
SIRT7, H3K18ac, and ELK4 Immunohistochemical Expression in Hepatocellular Carcinoma - Hye Seung Lee, Wonkyung Jung, Eunjung Lee1, Hyeyoon Chang, Jin Hyuk Choi, Han Gyeom Kim, Aeree Kim, Baek-hui Kim
-
Journal of Pathology and Translational Medicine 2016;50(5):337-344.
DOI: https://doi.org/10.4132/jptm.2016.05.20
Published online: August 5, 2016
Department of Pathology, Korea University Guro Hospital, Seoul, Korea
1Department of Pathology, Korea University Anam Hospital, Seoul, Korea
- Corresponding Author Baek-hui Kim, MD, PhD Department of Pathology, Korea University Guro Hospital, Korea University School of Medicine, 148 Gurodong-ro, Guro-gu, Seoul 08308, Korea Tel: +82-2-2626-1472 Fax: +82-2-2626-1486 E-mail: maelstrom@naver.com
© 2016 The Korean Society of Pathologists/The Korean Society for Cytopathology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted noncommercial use, distribution, and reproduction in any medium, provided the original work is properly cited.

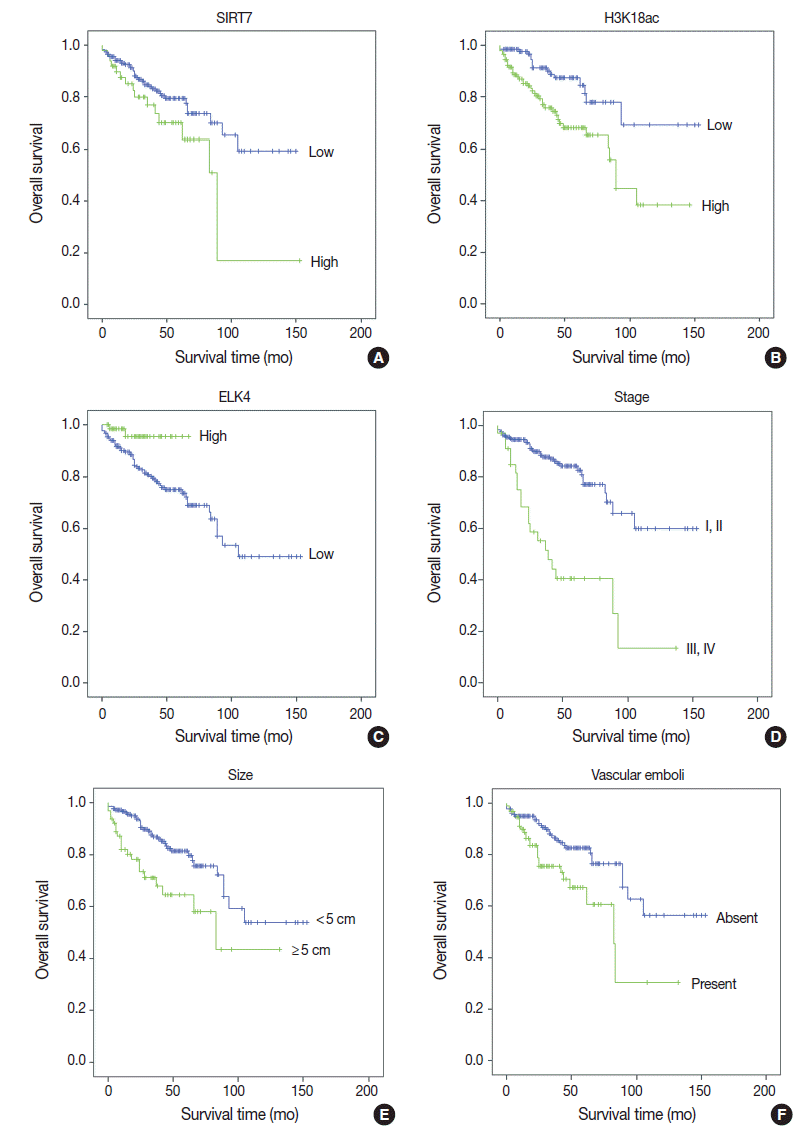
Figure & Data
References
Citations
- Combined single-cell RNA sequencing and mendelian randomization to identify biomarkers associated with necrotic apoptosis in intervertebral disc degeneration
Yi Ye, Lun Wan, Jiang Hu, Xiaoxue Li, Kun Zhang
The Spine Journal.2025; 25(1): 165. CrossRef - ETS-1 in tumor immunology: implications for novel anti-cancer strategies
SiYu Wang, Lei Wan, XiaoJun Zhang, HaoXiang Fang, MengYu Zhang, Feng Li, DaWei Yan
Frontiers in Immunology.2025;[Epub] CrossRef - Role of SIRT7 in Prostate Cancer Progression: New Insight Into Potential Therapeutic Target
Jiale Zhang, Chenxin Liu, Wenting Luo, Baoqing Sun
Cancer Medicine.2025;[Epub] CrossRef - SIRT7: the seventh key to unlocking the mystery of aging
Umar Raza, Xiaolong Tang, Zuojun Liu, Baohua Liu
Physiological Reviews.2024; 104(1): 253. CrossRef - The Significance of Modified Histone H3 in Epithelial Dysplasia and Oral Cancer
Woraphaluck Tachaveeraphong, Ekarat Phattarataratip
International Dental Journal.2024; 74(4): 769. CrossRef - Analysis of the Expression and Prognostic Value of SIRTs in Hepatocellular Carcinoma
Chuang Qin, Xiaofei Ye, Hongliang Luo, Hu Jin, Qiang Liu, Jiangfa Li
International Journal of General Medicine.2024; Volume 17: 2655. CrossRef - Role of Sirtuins in the Pathogenesis of Rheumatoid Arthritis
Agata Poniewierska-Baran, Oliwia Bochniak, Paulina Warias, Andrzej Pawlik
International Journal of Molecular Sciences.2023; 24(2): 1532. CrossRef - Role of sirtuins in hepatocellular carcinoma progression and multidrug resistance: Mechanistical and pharmacological perspectives
María Paula Ceballos, Ariel Darío Quiroga, Nicolás Francisco Palma
Biochemical Pharmacology.2023; 212: 115573. CrossRef - Substrates and Cyclic Peptide Inhibitors of the Oligonucleotide‐Activated Sirtuin 7**
Julie E. Bolding, Alexander L. Nielsen, Iben Jensen, Tobias N. Hansen, Line A. Ryberg, Samuel T. Jameson, Pernille Harris, Günther H. J. Peters, John M. Denu, Joseph M. Rogers, Christian A. Olsen
Angewandte Chemie International Edition.2023;[Epub] CrossRef - Substrates and Cyclic Peptide Inhibitors of the Oligonucleotide‐Activated Sirtuin 7**
Julie E. Bolding, Alexander L. Nielsen, Iben Jensen, Tobias N. Hansen, Line A. Ryberg, Samuel T. Jameson, Pernille Harris, Günther H. J. Peters, John M. Denu, Joseph M. Rogers, Christian A. Olsen
Angewandte Chemie.2023;[Epub] CrossRef - Epigenomic interplay in tumor heterogeneity: Potential of epidrugs as adjunct therapy
Suvasmita Rath, Diptesh Chakraborty, Jyotsnarani Pradhan, Mohammad Imran Khan, Jagneshwar Dandapat
Cytokine.2022; 157: 155967. CrossRef - Distinct histone H3 modification profiles correlate with aggressive characteristics of salivary gland neoplasms
Aroonwan Lam-Ubol, Ekarat Phattarataratip
Scientific Reports.2022;[Epub] CrossRef - Acetyl-CoA: An interplay between metabolism and epigenetics in cancer
Yang Hao, Qin Yi, Xu XiaoWu, Chen WeiBo, Zu GuangChen, Chen XueMin
Frontiers in Molecular Medicine.2022;[Epub] CrossRef - Sirtuins (SIRTs) As a Novel Target in Gastric Cancer
Agata Poniewierska-Baran, Paulina Warias, Katarzyna Zgutka
International Journal of Molecular Sciences.2022; 23(23): 15119. CrossRef - Novel oncogenes and tumor suppressor genes in hepatocellular carcinoma
Fang Wang, Peter Breslin S J, Wei Qiu
Liver Research.2021; 5(4): 195. CrossRef - Acute high folic acid treatment in SH-SY5Y cells with and without MTHFR function leads to gene expression changes in epigenetic modifying enzymes, changes in epigenetic marks, and changes in dendritic spine densities
Daniel F. Clark, Rachael Schmelz, Nicole Rogers, Nuri E. Smith, Kimberly R. Shorter, Lorenzo Chiariotti
PLOS ONE.2021; 16(1): e0245005. CrossRef - The E-Twenty-Six Family in Hepatocellular Carcinoma: Moving into the Spotlight
Tongyue Zhang, Danfei Liu, Yijun Wang, Mengyu Sun, Limin Xia
Frontiers in Oncology.2021;[Epub] CrossRef - Upregulation of histone acetylation reverses organic anion transporter 2 repression and enhances 5-fluorouracil sensitivity in hepatocellular carcinoma
Yingying Wang, Qianying Zhu, Haihong Hu, Hong Zhu, Bo Yang, Qiaojun He, Lushan Yu, Su Zeng
Biochemical Pharmacology.2021; 188: 114546. CrossRef - HCG11 up-regulation induced by ELK4 suppressed proliferation in vestibular schwannoma by targeting miR-620/ELK4
Ruiqing Long, Zhuohui Liu, Jinghui Li, Yuan Zhang, Hualin Yu
Cancer Cell International.2021;[Epub] CrossRef - Downregulation of circular RNA circPVT1 restricts cell growth of hepatocellular carcinoma through downregulation of Sirtuin 7 via microRNA‐3666
Yong Li, Haitao Shi, Jia Yuan, Lu Qiao, Lei Dong, Yan Wang
Clinical and Experimental Pharmacology and Physiology.2020; 47(7): 1291. CrossRef - Clinicopathological and molecular analysis of SIRT7 in hepatocellular carcinoma
Masae Yanai, Morito Kurata, Yutaka Muto, Hiroto Iha, Toshinori Kanao, Anna Tatsuzawa, Sachiko Ishibashi, Masumi Ikeda, Masanobu Kitagawa, Kouhei Yamamoto
Pathology.2020; 52(5): 529. CrossRef - MicroRNA‐148b Inhibits the Malignant Biological Behavior of Melanoma by Reducing Sirtuin 7 Expression Levels
Rui Sun, Meiliang Guo, Xiaojing Fan, Qinqin Meng, Dingfen Yuan, Xinrong Yang, Kexiang Yan, Hui Deng, Fengjie Sun
BioMed Research International.2020;[Epub] CrossRef - H3K18Ac as a Marker of Cancer Progression and Potential Target of Anti-Cancer Therapy
Marta Hałasa, Anna Wawruszak, Alicja Przybyszewska, Anna Jaruga, Małgorzata Guz, Joanna Kałafut, Andrzej Stepulak, Marek Cybulski
Cells.2019; 8(5): 485. CrossRef - Sirtuin7 has an oncogenic potential via promoting the growth of cholangiocarcinoma cells
Wenzhi Li, Zhe Sun, Chen Chen, Lin Wang, Zhimin Geng, Jie Tao
Biomedicine & Pharmacotherapy.2018; 100: 257. CrossRef - Identification of cancer‐related potential biomarkers based on lncRNA–pseudogene–mRNA competitive networks
Cheng Wu, Yunzhen Wei, Yinling Zhu, Kun Li, Yanjiao Zhu, Yichuan Zhao, Zhiqiang Chang, Yan Xu
FEBS Letters.2018; 592(6): 973. CrossRef - SIRT7 suppresses the epithelial-to-mesenchymal transition in oral squamous cell carcinoma metastasis by promoting SMAD4 deacetylation
Wenlu Li, Dandan Zhu, Shuaihua Qin
Journal of Experimental & Clinical Cancer Research.2018;[Epub] CrossRef - Sirtuin 7: a new marker of aggressiveness in prostate cancer
Romain Haider, Fabienne Massa, Lisa Kaminski, Stephan Clavel, Zied Djabari, Guillaume Robert, Kathiane Laurent, Jean-François Michiels, Matthieu Durand, Jean-Ehrland Ricci, Jean-François Tanti, Frédéric Bost, Damien Ambrosetti
Oncotarget.2017; 8(44): 77309. CrossRef
 PubReader
PubReader ePub Link
ePub Link-
 Cite this Article
Cite this Article
- Cite this Article
-
- Close
- Download Citation
- Close
- Figure
-
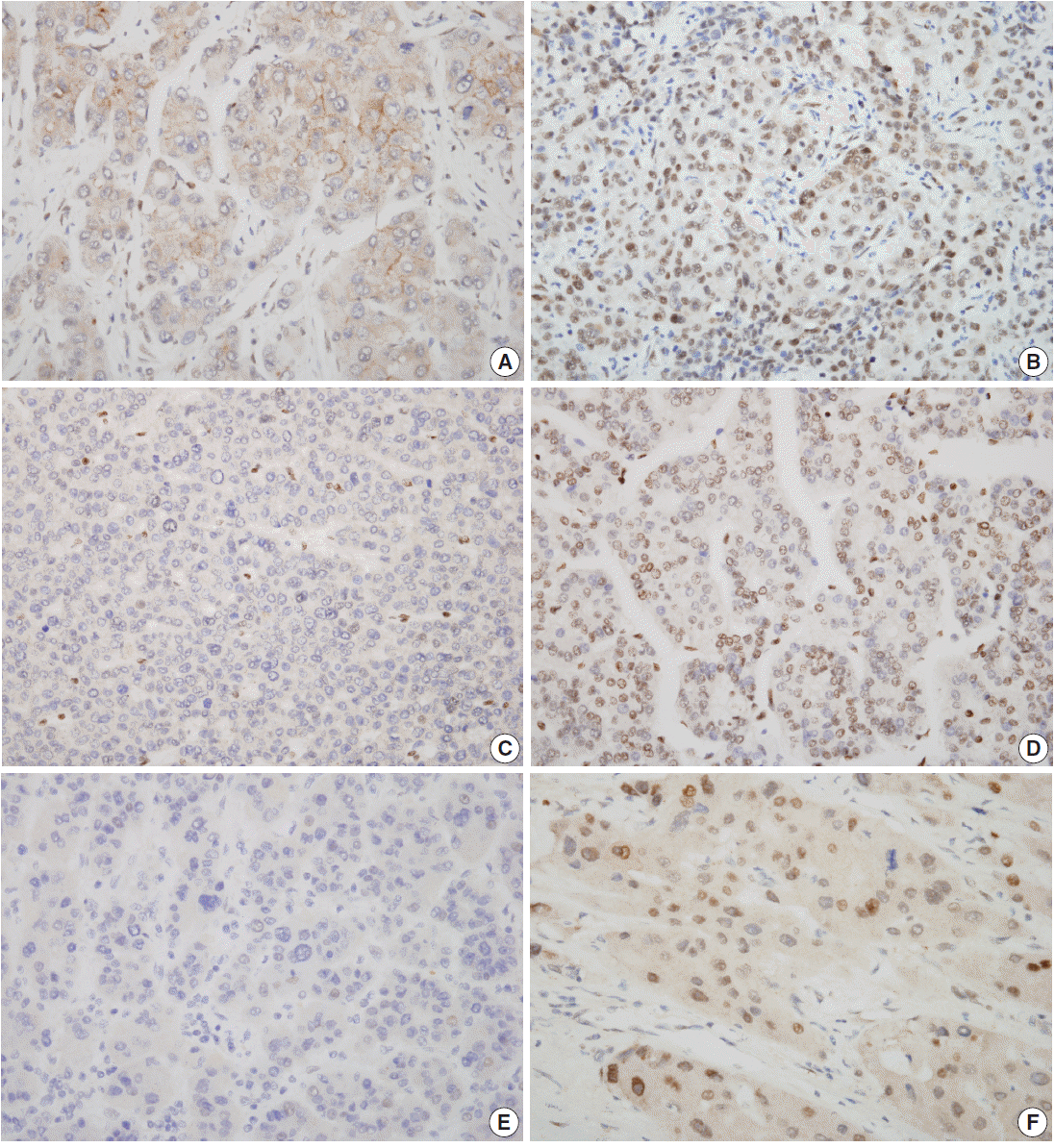
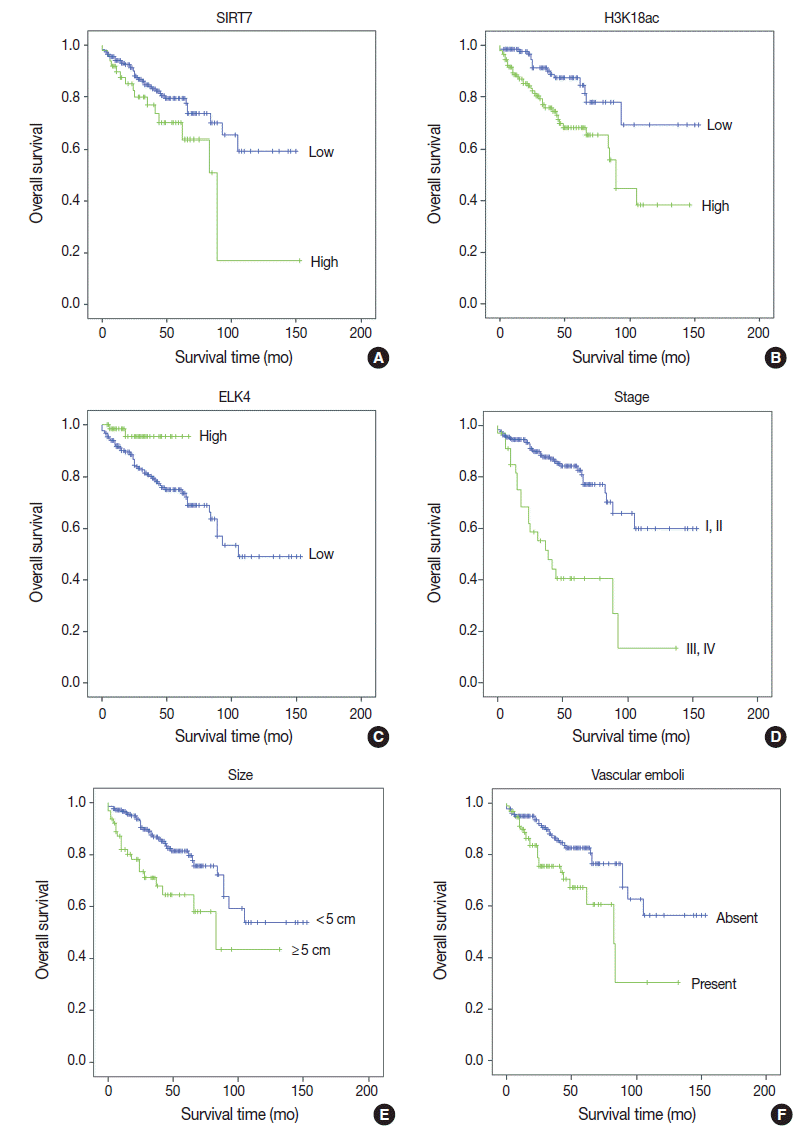


Fig. 1.
Fig. 2.
| Characteristic | No. (%) | Immunohistochemistry expression |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SIRT7 (n = 275) |
H3K18Ac (n = 276) |
ELK4 (n = 278) |
||||||||
| Low (n = 224, 81%) | High (n = 51, 19%) | p-value | Low (n = 133, 48%) | High (n = 143, 52%) | p-value | Low (n = 222, 76%) | High (n = 66, 24%) | p-value | ||
| Age (yr) | ||||||||||
| < 65 | 214 (77) | 176 | 37 | .721 | 80 | 115 | .184 | 80 | 115 | .184 |
| ≥ 65 | 64 (23) | 50 | 12 | 35 | 28 | 35 | 28 | |||
| Sex | ||||||||||
| Female | 47 (16.9) | 36 | 9 | .448 | 16 | 31 | .021 |
16 | 31 | .033 |
| Male | 231 (83) | 190 | 40 | 117 | 112 | 117 | 112 | |||
| Edmondson-Steiner grade | ||||||||||
| I–II | 178 (64) | 154 | 23 | .002 |
107 | 76 | < .001 |
135 | 43 | .828 |
| III–IV | 100 (36) | 72 | 26 | 26 | 67 | 77 | 23 | |||
| Tumor size (cm) | ||||||||||
| < 5 | 214 (77) | 176 | 37 | .721 | 103 | 111 | .972 | 163 | 51 | .948 |
| ≥ 5 | 64 (23) | 50 | 12 | 30 | 32 | 49 | 15 | |||
| AJCC stage | ||||||||||
| I–II | 244 (87.8) | 199 | 42 | .653 | 119 | 123 | .384 | 183 | 61 | .188 |
| III–IV | 34 (12.2) | 27 | 7 | 14 | 20 | 29 | 5 | |||
| Vascular emboli | ||||||||||
| Absent | 183 (65.8) | 153 | 29 | .255 | 98 | 85 | .012 |
148 | 35 | .056 |
| Present | 95 (34.2) | 73 | 20 | 35 | 58 | 64 | 29 | |||
| Margin | ||||||||||
| Negative | 263 (94.6) | 214 | 46 | .821 | 125 | 136 | .683 | 199 | 64 | .727 |
| Positive | 15 (5.4) | 12 | 3 | 8 | 7 | 13 | 2 | |||
| Tumor necrosis | ||||||||||
| Absent | 158 (61.7) | 131 | 25 | .261 | 80 | 77 | .133 | 113 | 45 | .211 |
| Present | 98 (38.3) | 76 | 21 | 40 | 57 | 77 | 21 | |||
| Variable | Univariate analysis |
Multivariate analysis |
||
|---|---|---|---|---|
| HR (95% CI) | p-value | HR (95% CI) | p-value | |
| High SIRT7 expression | 1.853 (1.036–3.313) | .038 |
1.323 (0.720–2.430) | .368 |
| High H3K18Ac expression | 2.569 (1.438–4.592) | .001 |
2.731 (1.469–5.076) | .001 |
| High ELK4 expression | 0.219 (0.053–0.907) | .036 |
0.162 (0.037–0.704) | .015 |
| Tumor size (≥ 5 cm) | 2.427 (1.400–4.207) | .002 |
2.044 (1.104–3.787) | .023 |
| Vascular emboli | 2.176 (1.273–3.720) | .004 |
1.287 (0.707–2.343) | .409 |
| Margin positive | 3.371 (1.582–7.183) | .002 |
2.943 (1.300–6.661) | .010 |
| AJCC stage III–IV | 4.302 (2.489–7.438) | < .001 |
3.007 (1.650–5.483) | < .001 |
| Variable | Univariate analysis |
Multivariate analysis |
||
|---|---|---|---|---|
| HR (95% CI) | p-value | HR (95% CI) | p-value | |
| High SIRT7 expression | 1.573 (1.013–2.444) | .044 |
1.440 (0.912–2.276) | .118 |
| High H3K18Ac expression | 1.229 (0.844–1.789) | .283 | 1.022 (0.680–1.537) | .915 |
| High ELK4 expression | 0.922 (0.566–1.502) | .744 | 0.901 (0.545–1.490) | .685 |
| Tumor size (≥ 5 cm) | 1.393 (0.903–2.150) | .134 | 1.110 (0.697–1.769) | .660 |
| Vascular emboli | 1.465 (1.001–2.144) | .049 |
1.192 (0.790–1.800) | .402 |
| Margin positive | 1.200 (0.558–2.580) | .640 | 0.903 (0.403–2.025) | .805 |
| AJCC stage III–IV | 2.128 (1.310–3.458) | .002 |
1.898 (1.101–3.273) | .021 |
| Edmondson-steiner’s grade | 1.513 (1.040–2.202) | .030 |
1.270 (0.839–1.923) | .258 |
| SIRT7 | H3K18ac | ELK4 | ||
|---|---|---|---|---|
| SIRT7 | Correlation coefficient | 1.000 | 0.165 | –0.054 |
| p-value | .006 |
.372 | ||
| H3K18ac | Correlation coefficient | 0.165 | 1.000 | –0.035 |
| p-value | .006 |
.560 | ||
| ELK4 | Correlation coefficient | –0.054 | –0.035 | 1.000 |
| p-value | .372 | .560 |
ELK4, ETS-like transcription factor 4; AJCC, American Joint Committee on Cancer. Statistically significant (p < .05).
HR, hazard ratio; CI, confidence interval; ELK4, ETS-like transcription factor 4; AJCC, American Joint Committee on Cancer. Statistically significant (p < .05).
HR, hazard ratio; CI, confidence interval; ELK4, ETS-like transcription factor 4; AJCC, American Joint Committee on Cancer. Statistically significant (p < .05).
ELK4, ETS-like transcription factor 4. Statistically significant (p < .05).

